

Epigenetic regulation during human cortical development: Seq-ing answers from the brain to the organoid
- PMID: 33915225
- PMCID: PMC8387070
- DOI: 10.1016/j.neuint.2021.105039
Epigenetic regulation during human cortical development: Seq-ing answers from the brain to the organoid
Abstract
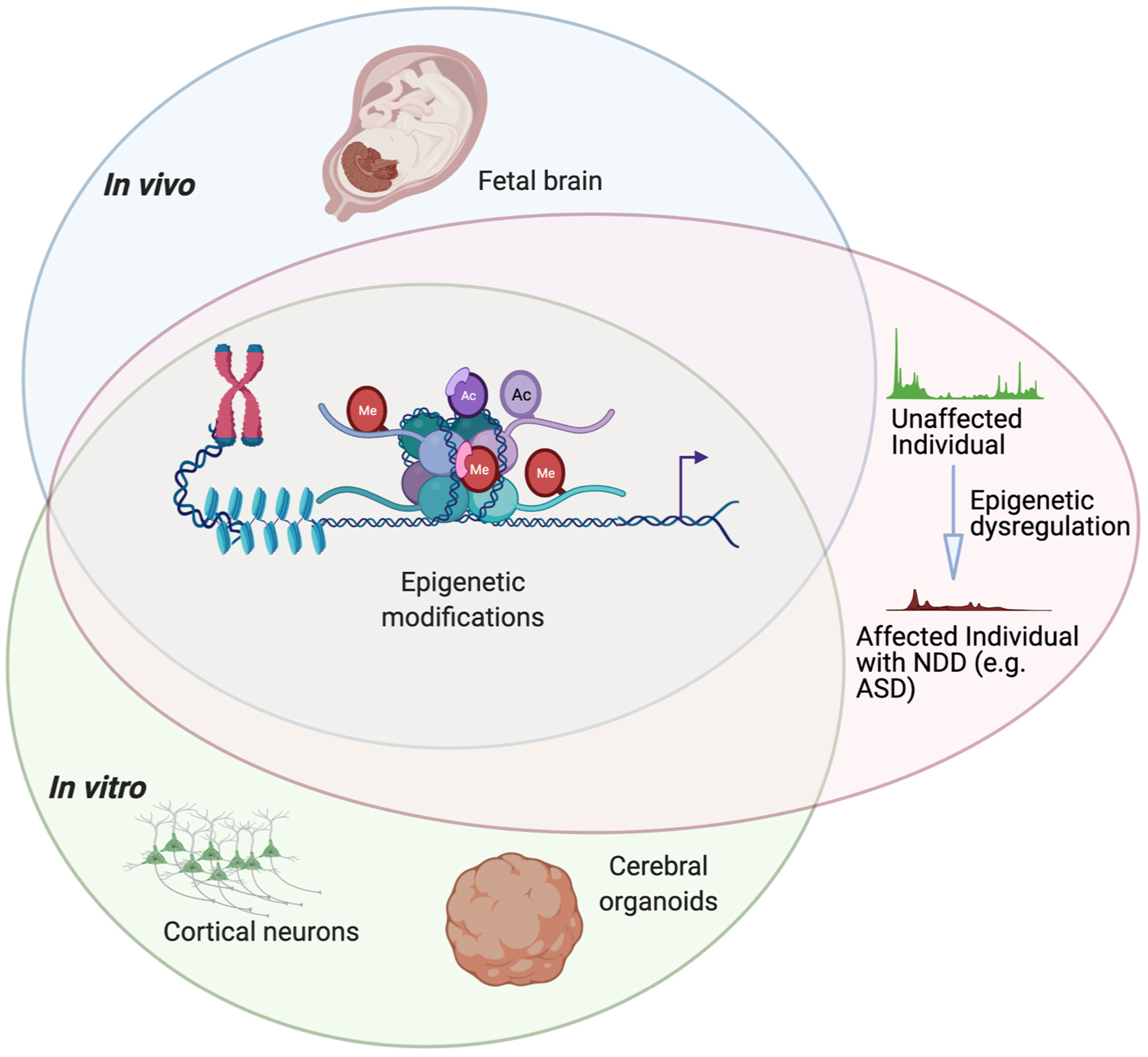
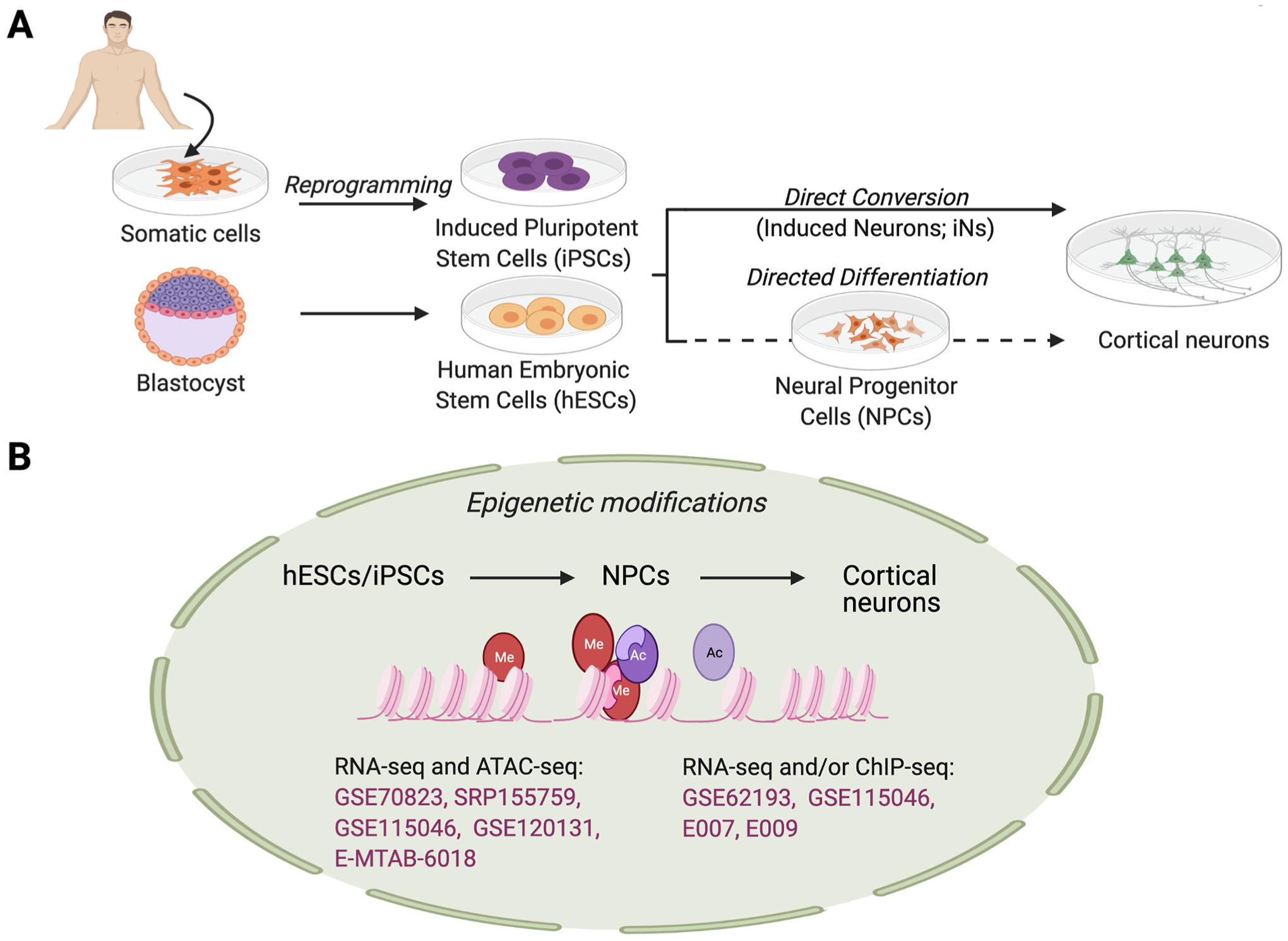
Epigenetic regulation plays an important role in controlling gene expression during complex processes, such as development of the human brain. Mutations in genes encoding chromatin modifying proteins and in the non-protein coding sequences of the genome can potentially alter transcription factor binding or chromatin accessibility. Such mutations can frequently cause neurodevelopmental disorders, therefore understanding how epigenetic regulation shapes brain development is of particular interest. While epigenetic regulation of neural development has been extensively studied in murine models, significant species-specific differences in both the genome sequence and in brain development necessitate human models. However, access to human fetal material is limited and these tissues cannot be grown or experimentally manipulated ex vivo. Therefore, models that recapitulate particular aspects of human fetal brain development, such as the in vitro differentiation of human pluripotent stem cells (hPSCs), are instrumental for studying the epigenetic regulation of human neural development. Here, we examine recent studies that have defined changes in the epigenomic landscape during fetal brain development. We compare these studies with analogous data derived by in vitro differentiation of hPSCs into specific neuronal cell types or as three-dimensional cerebral organoids. Such comparisons can be informative regarding which aspects of fetal brain development are faithfully recapitulated by in vitro differentiation models and provide a foundation for using experimentally tractable in vitro models of human brain development to study neural gene regulation and the basis of its disruption to cause neurodevelopmental disorders.
Keywords: Chromatin; Epigenetic regulation; Human brain development; Neuron; Organoid; Pluripotent stem cells.
Copyright © 2021 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of competing interests
Declarations of interest: none.
Figures


Similar articles
-
Cerebral Organoids Recapitulate Epigenomic Signatures of the Human Fetal Brain.Cell Rep. 2016 Dec 20;17(12):3369-3384. doi: 10.1016/j.celrep.2016.12.001. Cell Rep. 2016. PMID: 28009303 Free PMC article.
-
Mouse vs man: Organoid models of brain development & disease.Brain Res. 2019 Dec 1;1724:146427. doi: 10.1016/j.brainres.2019.146427. Epub 2019 Aug 29. Brain Res. 2019. PMID: 31473222 Review.
-
Comparative Transcriptomic Analysis of Cerebral Organoids and Cortical Neuron Cultures Derived from Human Induced Pluripotent Stem Cells.Stem Cells Dev. 2020 Nov 1;29(21):1370-1381. doi: 10.1089/scd.2020.0069. Epub 2020 Sep 22. Stem Cells Dev. 2020. PMID: 32862797 Free PMC article.
-
Dynamic Characterization of Structural, Molecular, and Electrophysiological Phenotypes of Human-Induced Pluripotent Stem Cell-Derived Cerebral Organoids, and Comparison with Fetal and Adult Gene Profiles.Cells. 2020 May 23;9(5):1301. doi: 10.3390/cells9051301. Cells. 2020. PMID: 32456176 Free PMC article.
-
Development and disease in a dish: the epigenetics of neurodevelopmental disorders.Epigenomics. 2018 Feb;10(2):219-231. doi: 10.2217/epi-2017-0113. Epub 2018 Jan 15. Epigenomics. 2018. PMID: 29334242 Free PMC article. Review.
Cited by
-
Effect of duty cycles of tumor‑treating fields on glioblastoma cells and normal brain organoids.Int J Oncol. 2022 Jan;60(1):8. doi: 10.3892/ijo.2021.5298. Epub 2021 Dec 31. Int J Oncol. 2022. PMID: 34970698 Free PMC article.
-
Transgenerational Effects of Prenatal Ethanol Exposure in Prepubescent Mice.Front Cell Dev Biol. 2022 Mar 21;10:812429. doi: 10.3389/fcell.2022.812429. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35386207 Free PMC article.
-
Brain organoid protocols and limitations.Front Cell Neurosci. 2024 Mar 20;18:1351734. doi: 10.3389/fncel.2024.1351734. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38572070 Free PMC article. Review.
-
AUTS2 Syndrome: Molecular Mechanisms and Model Systems.Front Mol Neurosci. 2022 Mar 31;15:858582. doi: 10.3389/fnmol.2022.858582. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35431798 Free PMC article. Review.
References
-
- Amiri A, Coppola G, Scuderi S, Wu F, Roychowdhury T, Liu F, Pochareddy S, Shin Y, Safi A, Song L, Zhu Y, Sousa AMM, Gerstein M, Crawford GE, Sestan N, Abyzov A, Vaccarino FM, 2018. Transcriptome and epigenome landscape of human cortical development modeled in brain organoids. Science 362. 10.1126/science.aat6720. - DOI - PMC - PubMed
-
- An J-Y, Lin K, Zhu L, Werling DM, Dong S, Brand H, Wang HZ, Zhao X, Schwartz GB, Collins RL, Currall BB, Dastmalchi C, Dea J, Duhn C, Gilson MC, Klei L, Liang L, Markenscoff-Papadimitriou E, Pochareddy S, Ahituv N, Buxbaum JD, Coon H, Daly MJ, Kim YS, Marth GT, Neale BM, Quinlan AR, Rubenstein JL, Sestan N, State MW, Willsey AJ, Talkowski ME, Devlin B, Roeder K, Sanders SJ, 2018. Genome-wide de novo risk score implicates promoter variation in autism spectrum disorder. Science 362. 10.1126/science.aat6576. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
