

MicroRNA-100 Mediates Hydrogen Peroxide-Induced Apoptosis of Human Retinal Pigment Epithelium ARPE-19 Cells
- PMID: 33915898
- PMCID: PMC8067261
- DOI: 10.3390/ph14040314
MicroRNA-100 Mediates Hydrogen Peroxide-Induced Apoptosis of Human Retinal Pigment Epithelium ARPE-19 Cells
Abstract
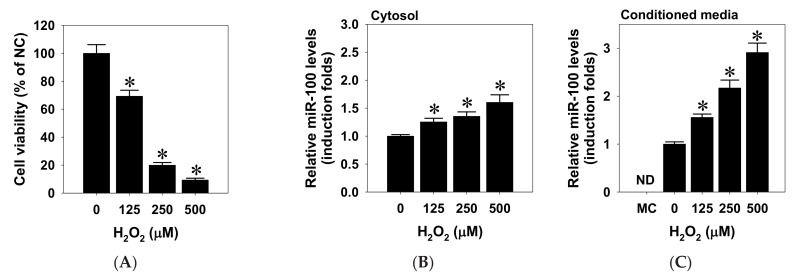
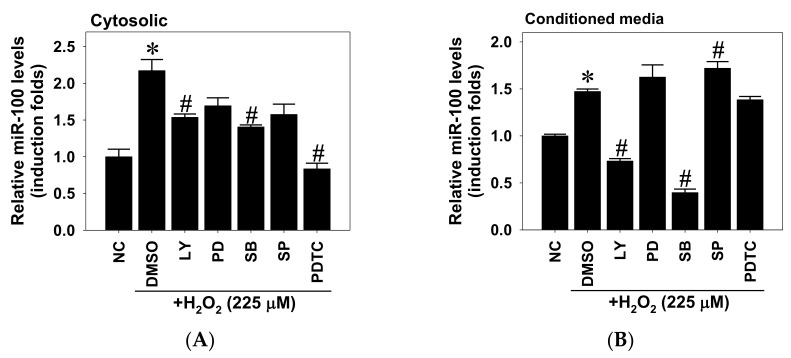
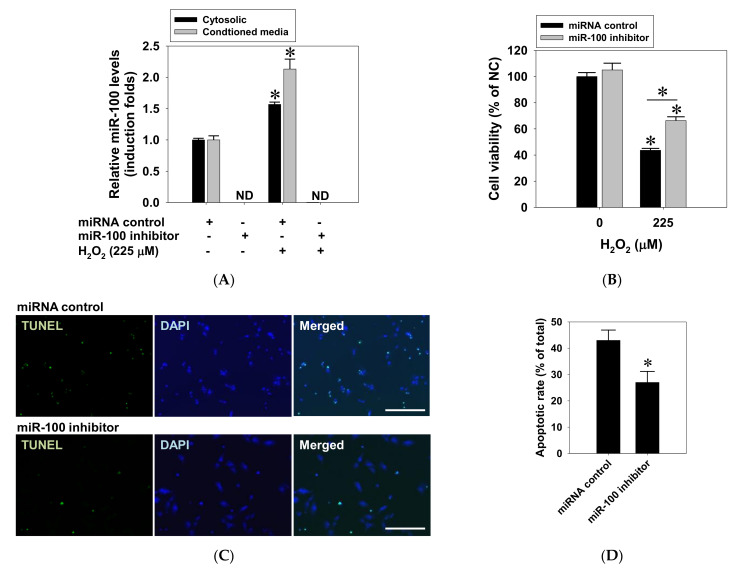
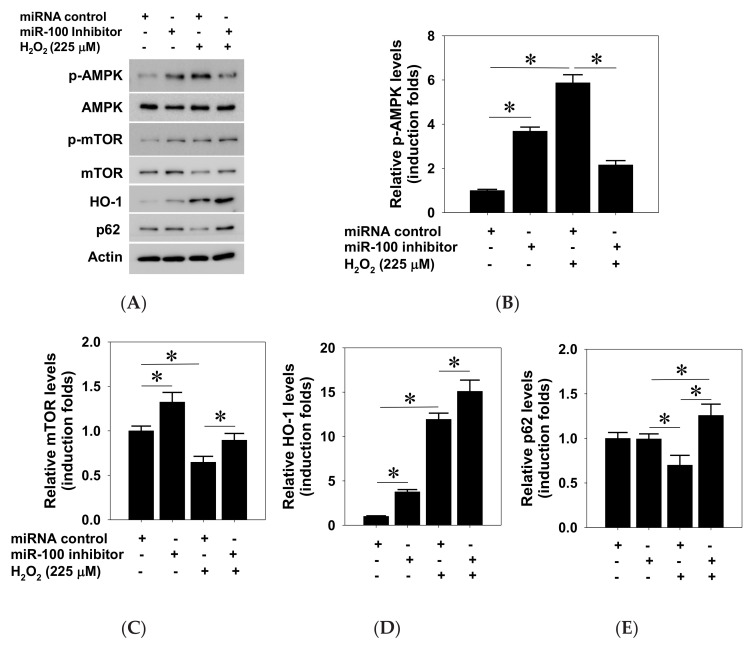
This study investigated the regulatory role of microRNA 100 (miR-100) in hydrogen peroxide (H2O2)-induced apoptosis of human retinal pigment epithelial ARPE-19 cells. H2O2 induced oxidative cell death of cultured ARPE-19 cells was measured by cytotoxicity assay. qRT-PCR was used to quantify cytosolic and extracellular contents of miR-100. Kinase and miR-100 inhibition treatments were applied to determine the regulatory signaling pathways involved in cell death regulation. H2O2 dose-dependently reduced viability of ARPE-19 cells and simultaneously upregulated miR-100 levels in both cytosolic and extracellular compartments. Western blotting detection indicated that H2O2 elicited hyperphosphorylation of PI3K/Akt, ERK1/2, JNK, p38 MAPK, and p65 NF-κB. Further kinase inhibition experiments demonstrated that PI3K, p38 MAPK, and NF-κB activities were involved in oxidative-stress-induced miR-100 upregulation in ARPE-19 cells, while blockade of PI3K, JNK, and NF-κB signaling significantly attenuated the oxidative cell death. Intriguingly, MiR-100 antagomir treatment exerted a cytoprotective effect against the H2O2-induced oxidative cell death through attenuating the oxidation-induced AMPK hyperphosphorylation, restoring cellular mTOR and p62/SQSTM1 levels and upregulating heme oxygenase-1 expression. These findings support that miR-100 at least in part mediates H2O2-induced cell death of ARPE-19 cells and can be regarded as a preventive and therapeutic target for retinal degenerative disease.
Keywords: heme oxygenase-1; mTOR; microRNA biosynthesis; oxidative stress; signal transduction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
Inhibition of the oxidative stress-induced miR-23a protects the human retinal pigment epithelium (RPE) cells from apoptosis through the upregulation of glutaminase and glutamine uptake.Mol Biol Rep. 2016 Oct;43(10):1079-87. doi: 10.1007/s11033-016-4041-8. Epub 2016 Jul 13. Mol Biol Rep. 2016. PMID: 27411920
-
The Protective Effect of Hispidin against Hydrogen Peroxide-Induced Oxidative Stress in ARPE-19 Cells via Nrf2 Signaling Pathway.Biomolecules. 2019 Aug 19;9(8):380. doi: 10.3390/biom9080380. Biomolecules. 2019. PMID: 31430968 Free PMC article.
-
Piceatannol Protects Human Retinal Pigment Epithelial Cells against Hydrogen Peroxide Induced Oxidative Stress and Apoptosis through Modulating PI3K/Akt Signaling Pathway.Nutrients. 2019 Jul 4;11(7):1515. doi: 10.3390/nu11071515. Nutrients. 2019. PMID: 31277394 Free PMC article.
-
Targeting cullin 3 by miR-601 activates Nrf2 signaling to protect retinal pigment epithelium cells from hydrogen peroxide.Biochem Biophys Res Commun. 2019 Aug 6;515(4):679-687. doi: 10.1016/j.bbrc.2019.05.171. Epub 2019 Jun 7. Biochem Biophys Res Commun. 2019. PMID: 31178131
-
Monounsaturated oleic acid modulates autophagy flux and upregulates angiogenic factor production in human retinal pigment epithelial ARPE-19 cells.Life Sci. 2020 Oct 15;259:118391. doi: 10.1016/j.lfs.2020.118391. Epub 2020 Sep 3. Life Sci. 2020. PMID: 32891610
Cited by
-
Oxidative Stress: A Suitable Therapeutic Target for Optic Nerve Diseases?Antioxidants (Basel). 2023 Jul 20;12(7):1465. doi: 10.3390/antiox12071465. Antioxidants (Basel). 2023. PMID: 37508003 Free PMC article. Review.
-
Immunomodulatory and Antioxidant Drugs in Glaucoma Treatment.Pharmaceuticals (Basel). 2023 Aug 22;16(9):1193. doi: 10.3390/ph16091193. Pharmaceuticals (Basel). 2023. PMID: 37765001 Free PMC article. Review.
-
miR‑100: A key tumor suppressor regulatory factor in human malignant tumors (Review).Int J Mol Med. 2025 Apr;55(4):67. doi: 10.3892/ijmm.2025.5508. Epub 2025 Feb 28. Int J Mol Med. 2025. PMID: 40017111 Free PMC article. Review.
-
Oxidative stress in the eye and its role in the pathophysiology of ocular diseases.Redox Biol. 2023 Dec;68:102967. doi: 10.1016/j.redox.2023.102967. Epub 2023 Nov 18. Redox Biol. 2023. PMID: 38006824 Free PMC article. Review.
-
Fabrication of nanozyme thixotropic anionic hydrogel for treating fungal keratitis by Dectin-1/p38 pathway.Appl Microbiol Biotechnol. 2025 Jun 26;109(1):153. doi: 10.1007/s00253-025-13529-8. Appl Microbiol Biotechnol. 2025. PMID: 40569312 Free PMC article.
References
-
- Friedman D.S., O’Colmain B.J., Munoz B., Tomany S.C., McCarty C., de Jong P.T., Nemesure B., Mitchell P., Kempen J. Prevalence of age-related macular degeneration in the United States. Arch. Ophthalmol. 2004;122:564–572. - PubMed
-
- Yang K., Liang Y.B., Gao L.Q., Peng Y., Shen R., Duan X.R., Friedman D.S., Sun L.P., Mitchell P., Wang N.L., et al. Prevalence of age-related macular degeneration in a rural Chinese population: The Handan Eye Study. Ophthalmology. 2011;118:1395–1401. doi: 10.1016/j.ophtha.2010.12.030. - DOI - PubMed
-
- Kawasaki R., Wang J.J., Ji G.J., Taylor B., Oizumi T., Daimon M., Kato T., Kawata S., Kayama T., Tano Y., et al. Prevalence and risk factors for age-related macular degeneration in an adult Japanese population: The Funagata study. Ophthalmology. 2008;115:1376–1381. doi: 10.1016/j.ophtha.2007.11.015. - DOI - PubMed
-
- Chen S.J., Cheng C.Y., Peng K.L., Li A.F., Hsu W.M., Liu J.H., Chou P. Prevalence and associated risk factors of age-related macular degeneration in an elderly Chinese population in Taiwan: The Shihpai Eye Study. Investig. Ophthalmol. Vis. Sci. 2008;49:3126–3133. doi: 10.1167/iovs.08-1803. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
