

Dual Role of TNF and LTα in Carcinogenesis as Implicated by Studies in Mice
- PMID: 33917839
- PMCID: PMC8068266
- DOI: 10.3390/cancers13081775
Dual Role of TNF and LTα in Carcinogenesis as Implicated by Studies in Mice
Abstract
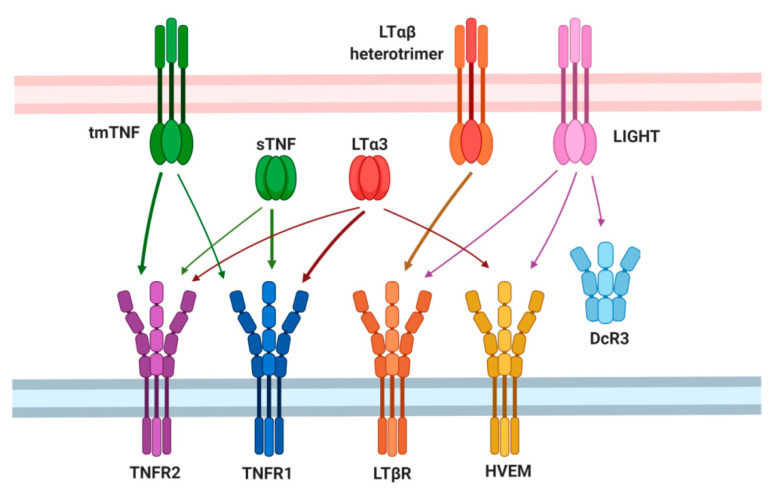
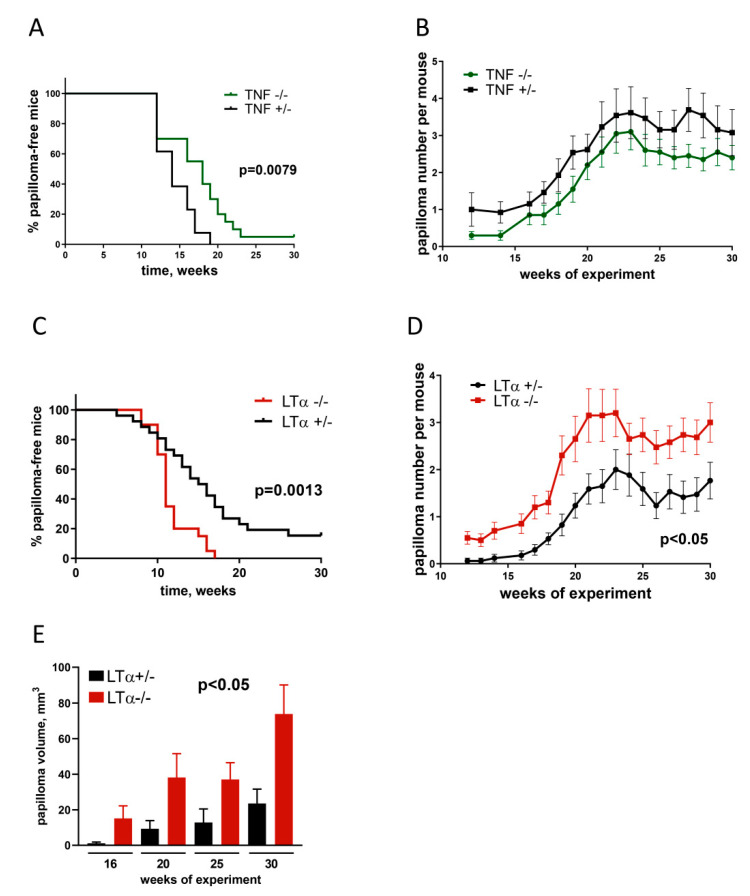
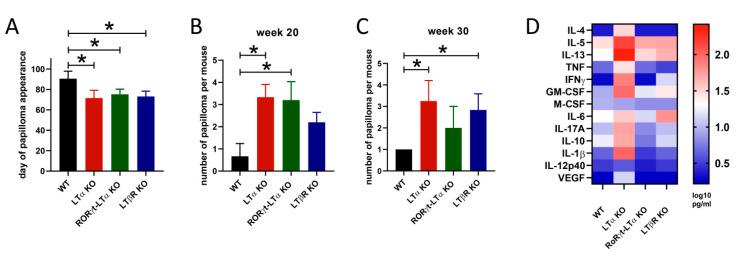
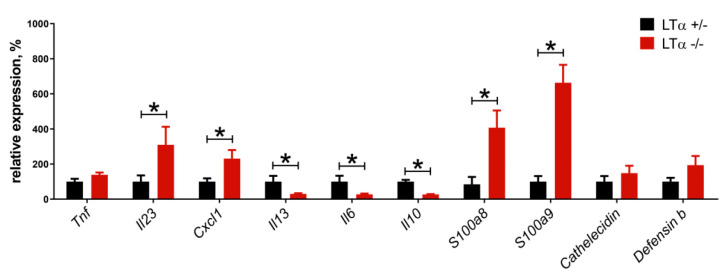
Tumor necrosis factor (TNF) and lymphotoxin alpha (LTα) are two related cytokines from the TNF superfamily, yet they mediate their functions in soluble and membrane-bound forms via overlapping, as well as distinct, molecular pathways. Their genes are encoded within the major histocompatibility complex class III cluster in close proximity to each other. TNF is involved in host defense, maintenance of lymphoid tissues, regulation of cell death and survival, and antiviral and antibacterial responses. LTα, known for some time as TNFβ, has pleiotropic functions including control of lymphoid tissue development and homeostasis cross talk between lymphocytes and their environment, as well as lymphoid tissue neogenesis with formation of lymphoid follicles outside the lymph nodes. Along with their homeostatic functions, deregulation of these two cytokines may be associated with initiation and progression of chronic inflammation, autoimmunity, and tumorigenesis. In this review, we summarize the current state of knowledge concerning TNF/LTα functions in tumor promotion and suppression, with the focus on the recently uncovered significance of host-microbiota interplay in cancer development that may explain some earlier controversial results.
Keywords: LTβR; TNFR2; cancer; lymphotoxin alpha; microbiota; mouse models; tumor necrosis factor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Distinct contributions of TNF and LT cytokines to the development of dendritic cells in vitro and their recruitment in vivo.Blood. 2003 Feb 15;101(4):1477-83. doi: 10.1182/blood.V101.4.1477. Blood. 2003. PMID: 12560241
-
Membrane lymphotoxin contributes to anti-leishmanial immunity by controlling structural integrity of lymphoid organs.Eur J Immunol. 2002 Jul;32(7):1993-2003. doi: 10.1002/1521-4141(200207)32:7<1993::AID-IMMU1993>3.0.CO;2-F. Eur J Immunol. 2002. PMID: 12115620
-
Independent protective effects for tumor necrosis factor and lymphotoxin alpha in the host response to Listeria monocytogenes infection.Infect Immun. 2005 Aug;73(8):4787-92. doi: 10.1128/IAI.73.8.4787-4792.2005. Infect Immun. 2005. PMID: 16040991 Free PMC article.
-
Tumor necrosis factor, lymphotoxin and cancer.IUBMB Life. 2010 Apr;62(4):283-9. doi: 10.1002/iub.309. IUBMB Life. 2010. PMID: 20155809 Review.
-
The unexpected role of lymphotoxin beta receptor signaling in carcinogenesis: from lymphoid tissue formation to liver and prostate cancer development.Oncogene. 2010 Sep 9;29(36):5006-18. doi: 10.1038/onc.2010.260. Epub 2010 Jul 5. Oncogene. 2010. PMID: 20603617 Review.
Cited by
-
Tumour Necrosis Factor Alpha (TNF-α) and Oral Squamous Cell Carcinoma.Cancers (Basel). 2023 Mar 19;15(6):1841. doi: 10.3390/cancers15061841. Cancers (Basel). 2023. PMID: 36980727 Free PMC article. Review.
-
Investigating Vitreous Cytokines in Choroidal Melanoma.Cancers (Basel). 2023 Jul 21;15(14):3701. doi: 10.3390/cancers15143701. Cancers (Basel). 2023. PMID: 37509362 Free PMC article.
-
Emerging Roles of Vitamin B12 in Aging and Inflammation.Int J Mol Sci. 2024 May 6;25(9):5044. doi: 10.3390/ijms25095044. Int J Mol Sci. 2024. PMID: 38732262 Free PMC article. Review.
-
The mechanism of cytokine regulation of cancer occurrence and development in the tumor microenvironment and its application in cancer treatment: a narrative review.Transl Cancer Res. 2024 Oct 31;13(10):5649-5663. doi: 10.21037/tcr-24-679. Epub 2024 Oct 18. Transl Cancer Res. 2024. PMID: 39525000 Free PMC article. Review.
-
Searching for New Biomarkers of Neuroendocrine Tumors: A Comparative Analysis of Chromogranin A and Inflammatory Cytokines in Patients with Neuroendocrine Tumors.Curr Oncol. 2024 Oct 12;31(10):6110-6132. doi: 10.3390/curroncol31100456. Curr Oncol. 2024. PMID: 39451760 Free PMC article.
References
-
- Browning J.L., Dougas I., Ngam-ek A., Bourdon P.R., Ehrenfels B.N., Miatkowski K., Zafari M., Yampaglia A.M., Lawton P., Meier W., et al. Characterization of surface lymphotoxin forms. Use of specific monoclonal antibodies and soluble receptors. J. Immunol. 1995;154:33–46. - PubMed
-
- Browning J.L., Ngam-ek A., Lawton P., DeMarinis J., Tizard R., Chow E.P., Hession C., O’Brine-Greco B., Foley S.F., Ware C.F. Lymphotoxin beta, a novel member of the TNF family that forms a heteromeric complex with lymphotoxin on the cell surface. Cell. 1993;72:847–856. doi: 10.1016/0092-8674(93)90574-A. - DOI - PubMed
-
- Pasparakis M., Alexopoulou L., Episkopou V., Kollias G. Immune and inflammatory responses in TNF alpha-deficient mice: A critical requirement for TNF alpha in the formation of primary B cell follicles, follicular dendritic cell networks and germinal centers, and in the maturation of the humoral immune response. J. Exp. Med. 1996;184:1397–1411. doi: 10.1084/jem.184.4.1397. - DOI - PMC - PubMed
-
- Korner H., Cook M., Riminton D.S., Lemckert F.A., Hoek R.M., Ledermann B., Kontgen F., Fazekas de St Groth B., Sedgwick J.D. Distinct roles for lymphotoxin-alpha and tumor necrosis factor in organogenesis and spatial organization of lymphoid tissue. Eur. J. Immunol. 1997;27:2600–2609. doi: 10.1002/eji.1830271020. - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
