

Glutathione Metabolism Contributes to the Induction of Trained Immunity
- PMID: 33919212
- PMCID: PMC8143087
- DOI: 10.3390/cells10050971
Glutathione Metabolism Contributes to the Induction of Trained Immunity
Abstract
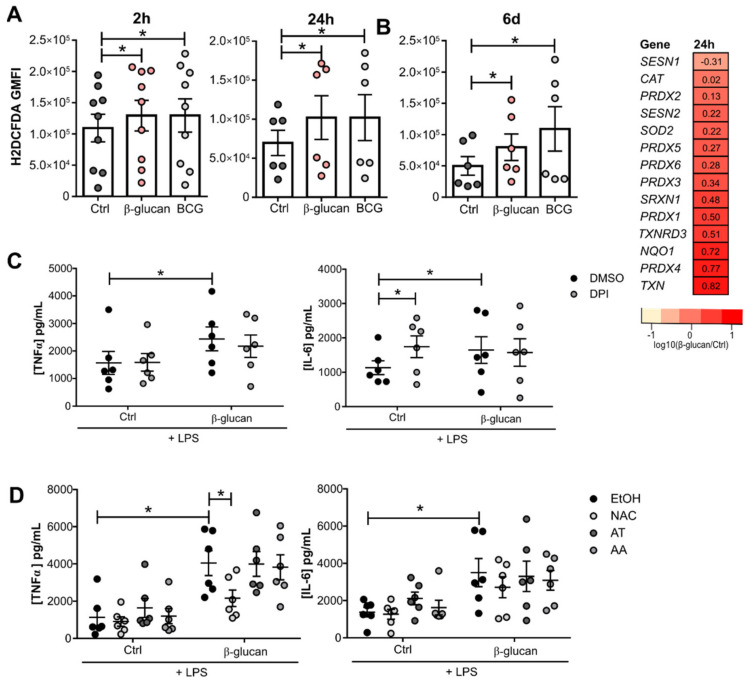
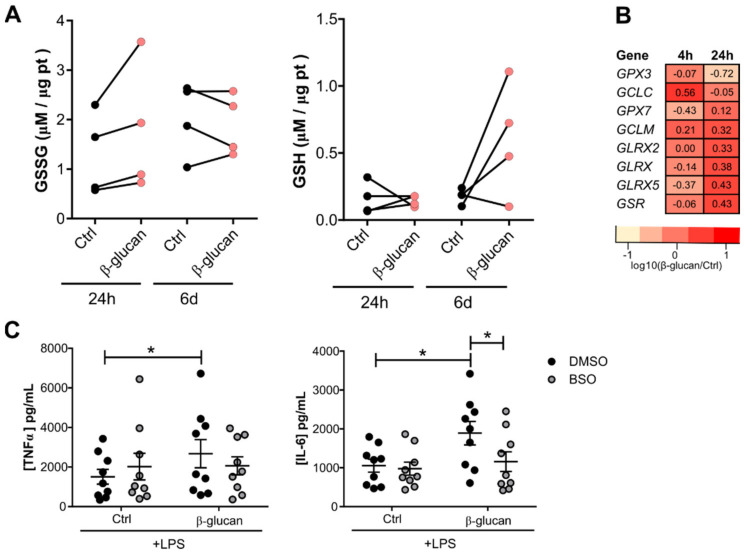
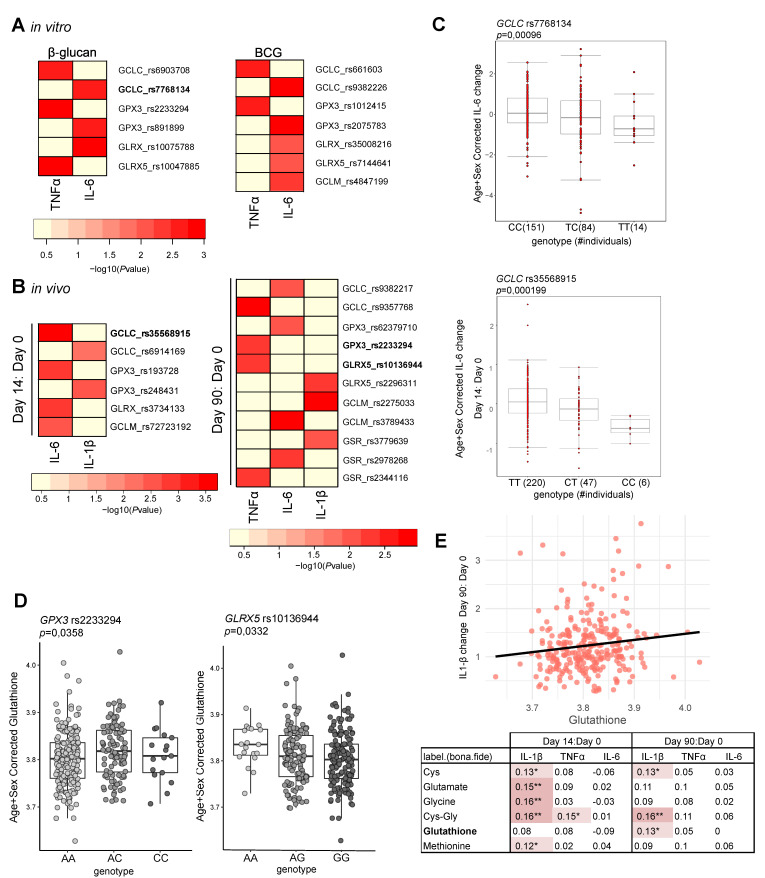
The innate immune system displays heterologous memory characteristics, which are characterized by stronger responses to a secondary challenge. This phenomenon termed trained immunity relies on epigenetic and metabolic rewiring of innate immune cells. As reactive oxygen species (ROS) production has been associated with the trained immunity phenotype, we hypothesized that the increased ROS levels and the main intracellular redox molecule glutathione play a role in the induction of trained immunity. Here we show that pharmacological inhibition of ROS in an in vitro model of trained immunity did not influence cell responsiveness; the modulation of glutathione levels reduced pro-inflammatory cytokine production in human monocytes. Single nucleotide polymorphisms (SNPs) in genes involved in glutathione metabolism were found to be associated with changes in pro-inflammatory cytokine production capacity upon trained immunity. Also, plasma glutathione concentrations were positively associated with ex vivo IL-1β production, a biomarker of trained immunity, produced by monocytes of BCG-vaccinated individuals. In conclusion, glutathione metabolism is involved in the induction of trained immunity, and future studies are warranted to explore its functional consequences in human diseases.
Keywords: glutathione; innate immune memory; macrophages; metabolism; trained immunity.
Conflict of interest statement
MGN is a scientific founder of TTxD. The remaining authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
Similar articles
-
Long-term histone lactylation connects metabolic and epigenetic rewiring in innate immune memory.Cell. 2025 May 29;188(11):2992-3012.e16. doi: 10.1016/j.cell.2025.03.048. Epub 2025 May 2. Cell. 2025. PMID: 40318634
-
Aldosterone induces trained immunity: the role of fatty acid synthesis.Cardiovasc Res. 2020 Feb 1;116(2):317-328. doi: 10.1093/cvr/cvz137. Cardiovasc Res. 2020. PMID: 31119285
-
In Vitro Experimental Model of Trained Innate Immunity in Human Primary Monocytes.Clin Vaccine Immunol. 2016 Dec 5;23(12):926-933. doi: 10.1128/CVI.00349-16. Print 2016 Dec. Clin Vaccine Immunol. 2016. PMID: 27733422 Free PMC article.
-
The Intersection of Epigenetics and Metabolism in Trained Immunity.Immunity. 2021 Jan 12;54(1):32-43. doi: 10.1016/j.immuni.2020.10.011. Epub 2020 Nov 20. Immunity. 2021. PMID: 33220235 Review.
-
Trained Innate Immunity as a Novel Mechanism Linking Infection and the Development of Atherosclerosis.Circ Res. 2018 Mar 2;122(5):664-669. doi: 10.1161/CIRCRESAHA.117.312465. Epub 2018 Jan 24. Circ Res. 2018. PMID: 29367213 Review.
Cited by
-
Regulation of immune cell function by nicotinamide nucleotide transhydrogenase.Am J Physiol Cell Physiol. 2022 Apr 1;322(4):C666-C673. doi: 10.1152/ajpcell.00607.2020. Epub 2022 Feb 9. Am J Physiol Cell Physiol. 2022. PMID: 35138175 Free PMC article. Review.
-
Nano-Drug Design Based on the Physiological Properties of Glutathione.Molecules. 2021 Sep 13;26(18):5567. doi: 10.3390/molecules26185567. Molecules. 2021. PMID: 34577040 Free PMC article. Review.
-
Training vs. Tolerance: The Yin/Yang of the Innate Immune System.Biomedicines. 2023 Mar 2;11(3):766. doi: 10.3390/biomedicines11030766. Biomedicines. 2023. PMID: 36979747 Free PMC article. Review.
-
Metabolomics in aging research: aging markers from organs.Front Cell Dev Biol. 2023 Jun 16;11:1198794. doi: 10.3389/fcell.2023.1198794. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37397261 Free PMC article. Review.
-
Dectin-1 Signaling Update: New Perspectives for Trained Immunity.Front Immunol. 2022 Feb 14;13:812148. doi: 10.3389/fimmu.2022.812148. eCollection 2022. Front Immunol. 2022. PMID: 35237264 Free PMC article. Review.
References
-
- Netea M.G., Domínguez-Andrés J., Barreiro L.B., Chavakis T., Divangahi M., Fuchs E., Joosten L.A.B., van der Meer J.W.M., Mhlanga M.M., Mulder W.J.M., et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020;20:375–388. doi: 10.1038/s41577-020-0285-6. - DOI - PMC - PubMed
-
- Quintin J., Saeed S., Martens J.H.A., Giamarellos-Bourboulis E.J., Ifrim D.C., Logie C., Jacobs L., Jansen T., Kullberg B.J., Wijmenga C., et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe. 2012;12:223–232. doi: 10.1016/j.chom.2012.06.006. - DOI - PMC - PubMed
-
- Bekkering S., Quintin J., Joosten L.A., Van Der Meer J.W., Netea M.G., Riksen N.P. Oxidized Low-Density Lipoprotein Induces Long-Term Proinflammatory Cytokine Production and Foam Cell Formation via Epigenetic Reprogramming of Monocytes. Arter. Thromb. Vasc. Biol. 2014;34:1731–1738. doi: 10.1161/ATVBAHA.114.303887. - DOI - PubMed
-
- Arts R.J.W., Novakovic B., ter Horst R., Carvalho A., Bekkering S., Lachmandas E., Rodrigues F., Silvestre R., Cheng S.C., Wang S.Y., et al. Glutaminolysis and Fumarate Accumulation Integrate Immunometabolic and Epigenetic Programs in Trained Immunity. Cell Metab. 2016;24:807–819. doi: 10.1016/j.cmet.2016.10.008. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
