

The Histone Chaperone HIRA Is a Positive Regulator of Seed Germination
- PMID: 33919775
- PMCID: PMC8070706
- DOI: 10.3390/ijms22084031
The Histone Chaperone HIRA Is a Positive Regulator of Seed Germination
Abstract
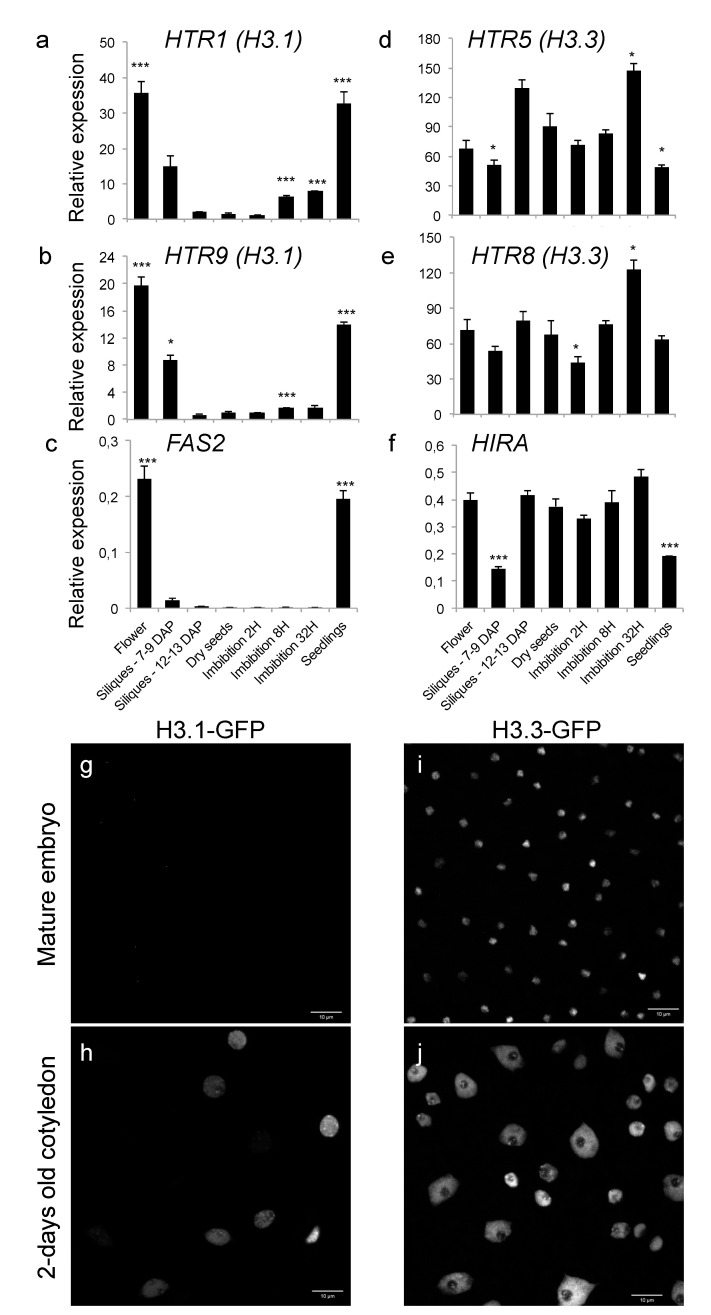
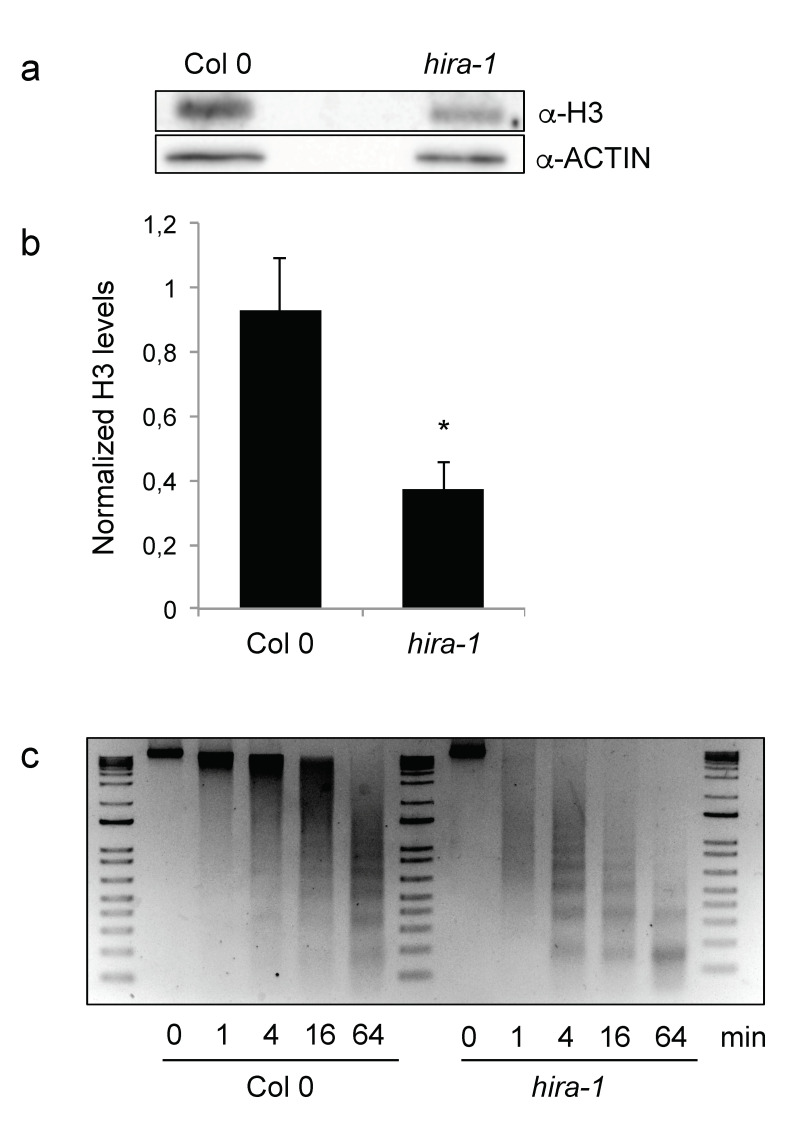
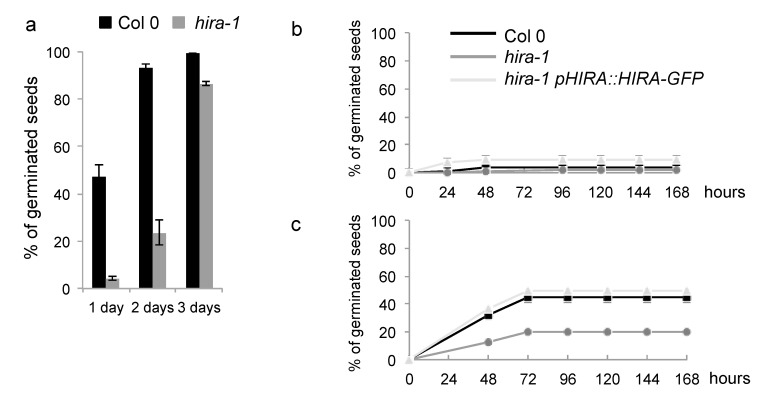
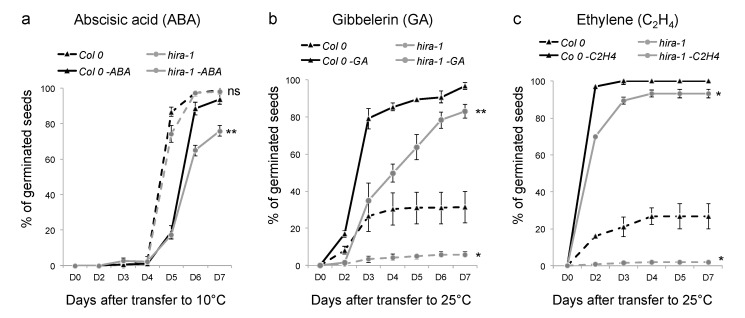
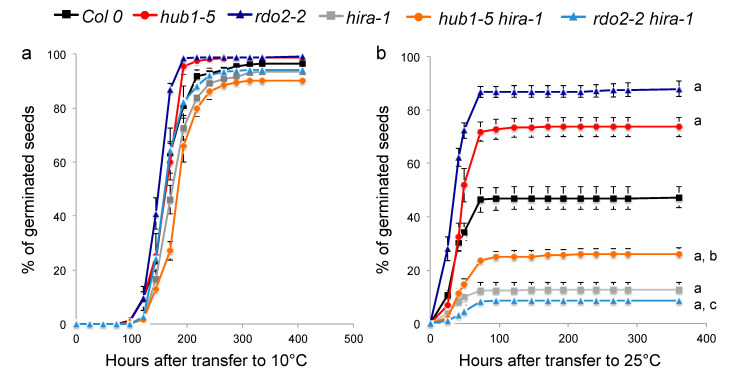
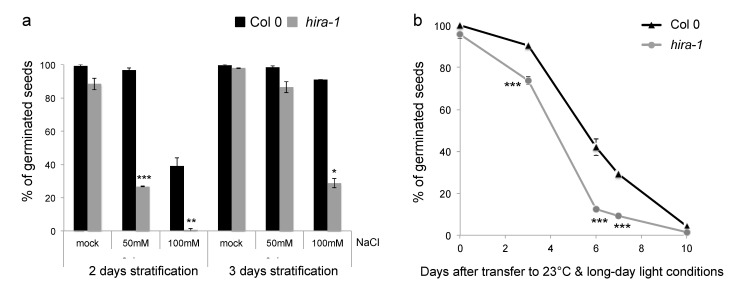
Histone chaperones regulate the flow and dynamics of histone variants and ensure their assembly into nucleosomal structures, thereby contributing to the repertoire of histone variants in specialized cells or tissues. To date, not much is known on the distribution of histone variants and their modifications in the dry seed embryo. Here, we bring evidence that genes encoding the replacement histone variant H3.3 are expressed in Arabidopsis dry seeds and that embryo chromatin is characterized by a low H3.1/H3.3 ratio. Loss of HISTONE REGULATOR A (HIRA), a histone chaperone responsible for H3.3 deposition, reduces cellular H3 levels and increases chromatin accessibility in dry seeds. These molecular differences are accompanied by increased seed dormancy in hira-1 mutant seeds. The loss of HIRA negatively affects seed germination even in the absence of HISTONE MONOUBIQUITINATION 1 or TRANSCRIPTION ELONGATION FACTOR II S, known to be required for seed dormancy. Finally, hira-1 mutant seeds show lower germination efficiency when aged under controlled deterioration conditions or when facing unfavorable environmental conditions such as high salinity. Altogether, our results reveal a dependency of dry seed chromatin organization on the replication-independent histone deposition pathway and show that HIRA contributes to modulating seed dormancy and vigor.
Keywords: chromatin; histone chaperones; histone variants; seed dormancy and germination; seed vigor.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
