

The Relationship between Oscillations in Brain Regions and Functional Connectivity: A Critical Analysis with the Aid of Neural Mass Models
- PMID: 33921414
- PMCID: PMC8069852
- DOI: 10.3390/brainsci11040487
The Relationship between Oscillations in Brain Regions and Functional Connectivity: A Critical Analysis with the Aid of Neural Mass Models
Abstract
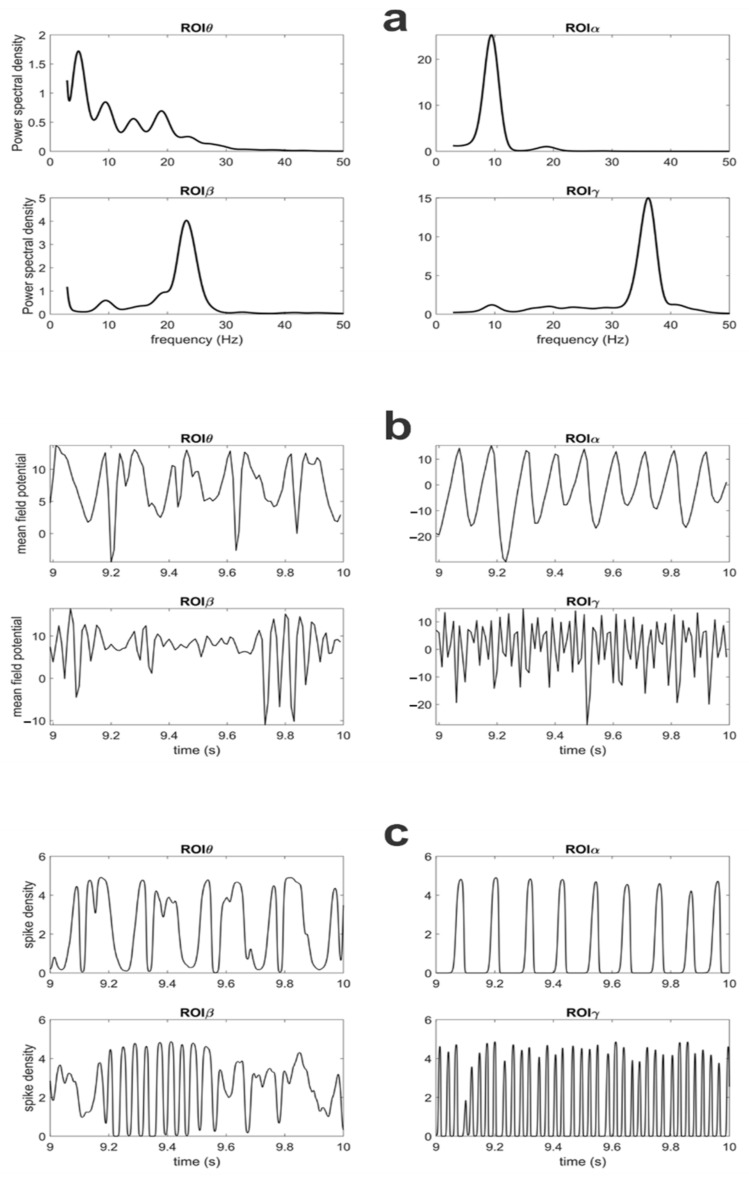
Propagation of brain rhythms among cortical regions is a relevant aspect of cognitive neuroscience, which is often investigated using functional connectivity (FC) estimation techniques. The aim of this work is to assess the relationship between rhythm propagation, FC and brain functioning using data generated from neural mass models of connected Regions of Interest (ROIs). We simulated networks of four interconnected ROIs, each with a different intrinsic rhythm (in θ, α, β and γ ranges). Connectivity was estimated using eight estimators and the relationship between structural connectivity and FC was assessed as a function of the connectivity strength and of the inputs to the ROIs. Results show that the Granger estimation provides the best accuracy, with a good capacity to evaluate the connectivity strength. However, the estimated values strongly depend on the input to the ROIs and hence on nonlinear phenomena. When a population works in the linear region, its capacity to transmit a rhythm increases drastically. Conversely, when it saturates, oscillatory activity becomes strongly affected by rhythms incoming from other regions. Changes in functional connectivity do not always reflect a physical change in the synapses. A unique connectivity network can propagate rhythms in very different ways depending on the specific working conditions.
Keywords: Granger causality; connectivity; cortical rhythms; excitatory and inhibitory synapses; neural mass models; nonlinear neural phenomena.
Conflict of interest statement
The authors declare no conflict of interest.
Figures












References
-
- Reid A.T., Headley D.B., Mill R.D., Sanchez-Romero R., Uddin L.Q., Marinazzo D., Lurie D.J., Valdés-Sosa P.A., Hanson S.J., Biswal B.B., et al. Advancing Functional Connectivity Research from Association to Causation. Nat. Neurosci. 2019;22:1751–1760. doi: 10.1038/s41593-019-0510-4. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
