

Evaluation of Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus Mosquitoes Competence to Oropouche virus Infection
- PMID: 33923055
- PMCID: PMC8145018
- DOI: 10.3390/v13050755
Evaluation of Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus Mosquitoes Competence to Oropouche virus Infection
Abstract
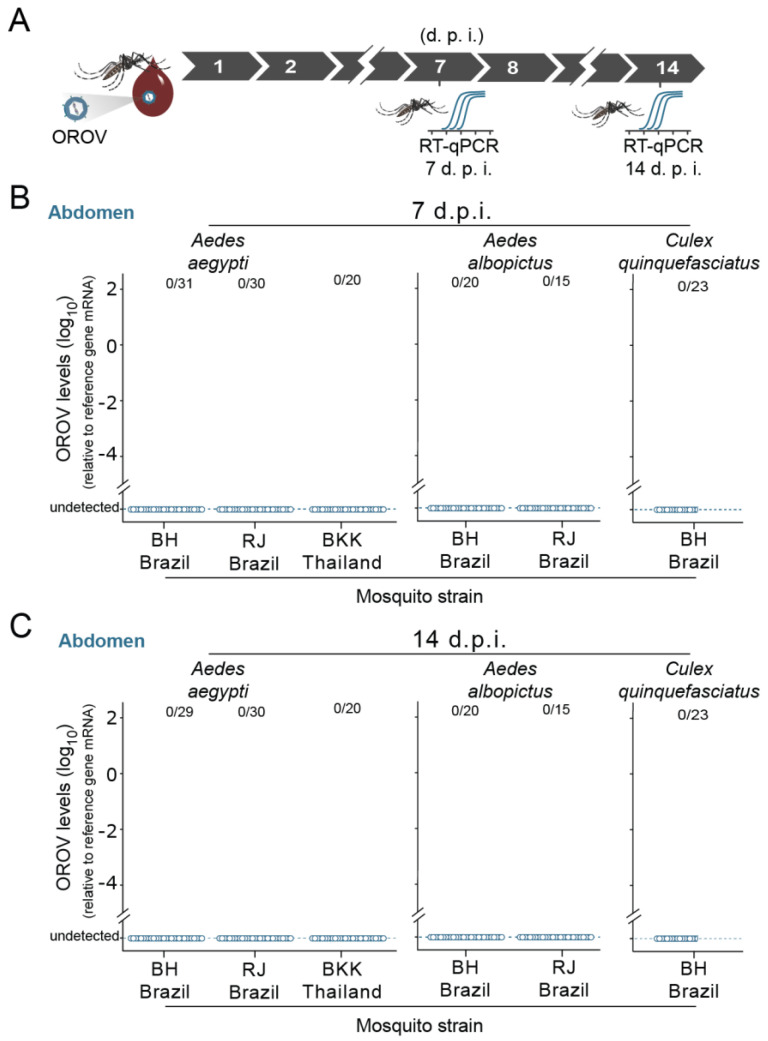
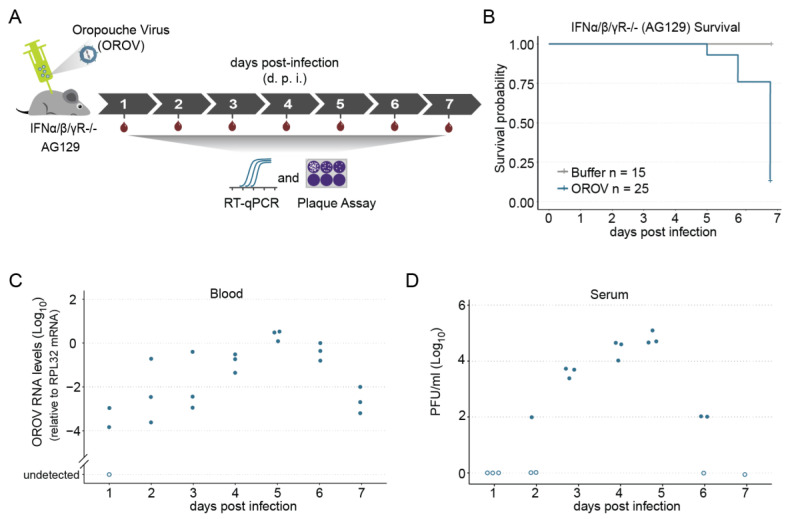
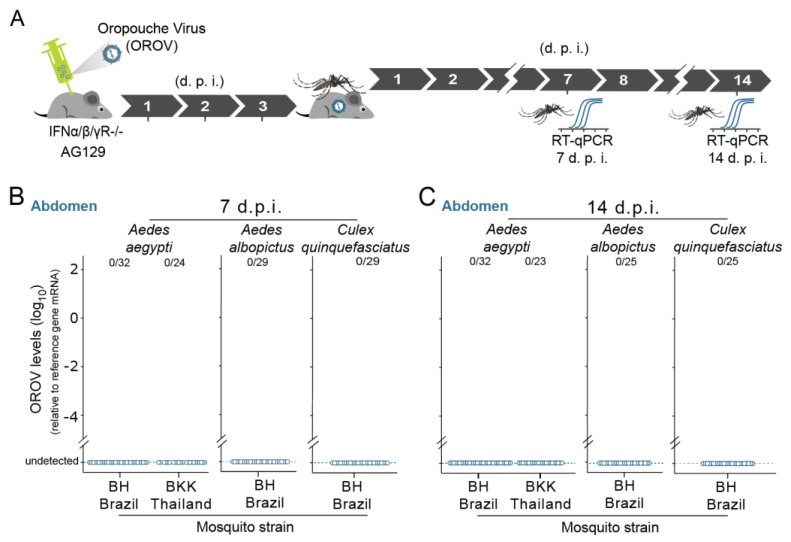
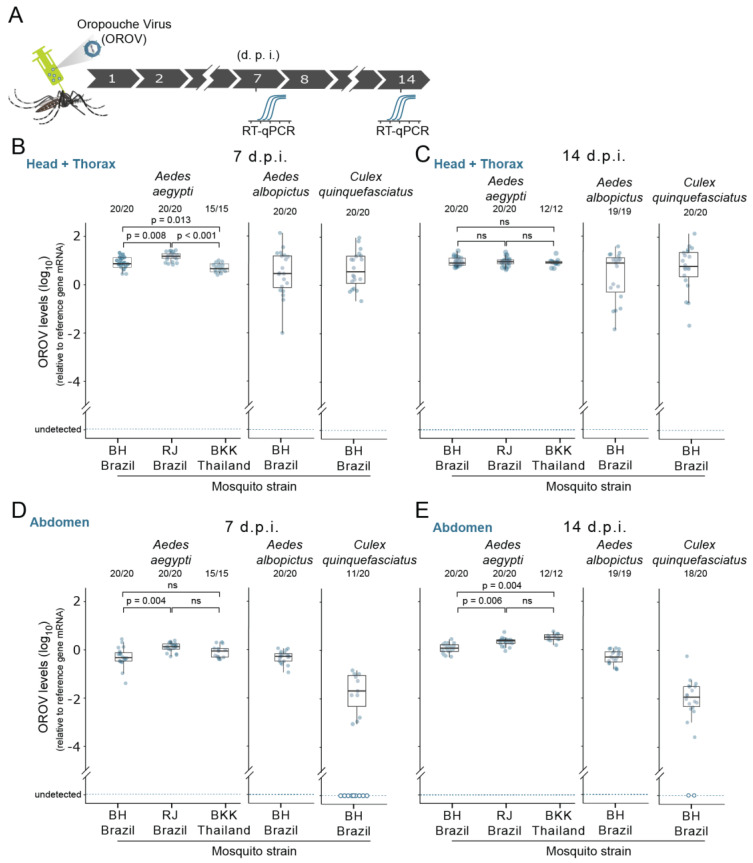
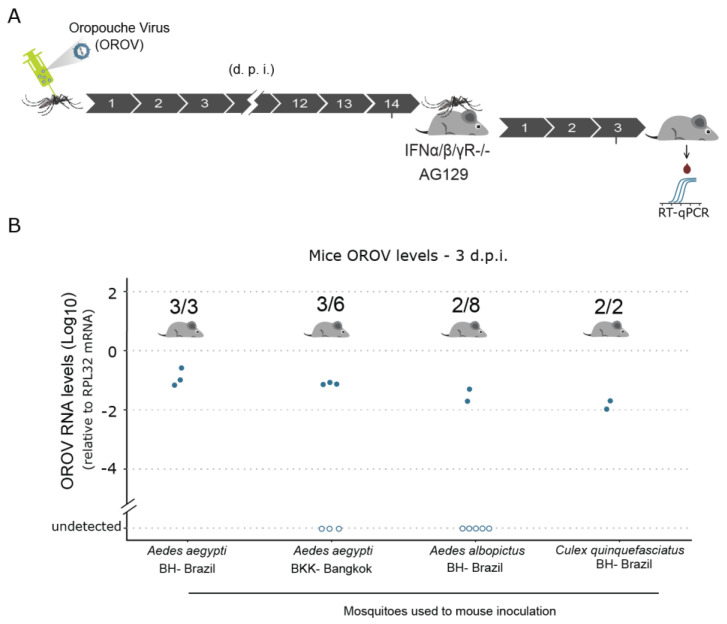
The emergence of new human viral pathogens and re-emergence of several diseases are of particular concern in the last decades. Oropouche orthobunyavirus (OROV) is an arbovirus endemic to South and Central America tropical regions, responsible to several epidemic events in the last decades. There is little information regarding the ability of OROV to be transmitted by urban/peri-urban mosquitoes, which has limited the predictability of the emergence of permanent urban transmission cycles. Here, we evaluated the ability of OROV to infect, replicate, and be transmitted by three anthropophilic and urban species of mosquitoes, Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus. We show that OROV is able to infect and efficiently replicate when systemically injected in all three species tested, but not when orally ingested. Moreover, we find that, once OROV replication has occurred in the mosquito body, all three species were able to transmit the virus to immunocompromised mice during blood feeding. These data provide evidence that OROV is restricted by the midgut barrier of three major urban mosquito species, but, if this restriction is overcome, could be efficiently transmitted to vertebrate hosts. This poses a great risk for the emergence of permanent urban cycles and geographic expansion of OROV to other continents.
Keywords: Aedes aegypti; Aedes albopictus; Culex quinquefasciatus; Oropouche; urban epidemics; vector competence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
