

Species-Specific Regulation of TRPM2 by PI(4,5)P2 via the Membrane Interfacial Cavity
- PMID: 33924946
- PMCID: PMC8125603
- DOI: 10.3390/ijms22094637
Species-Specific Regulation of TRPM2 by PI(4,5)P2 via the Membrane Interfacial Cavity
Abstract
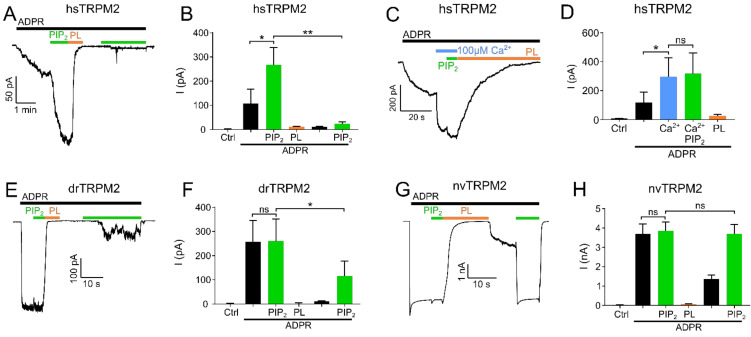
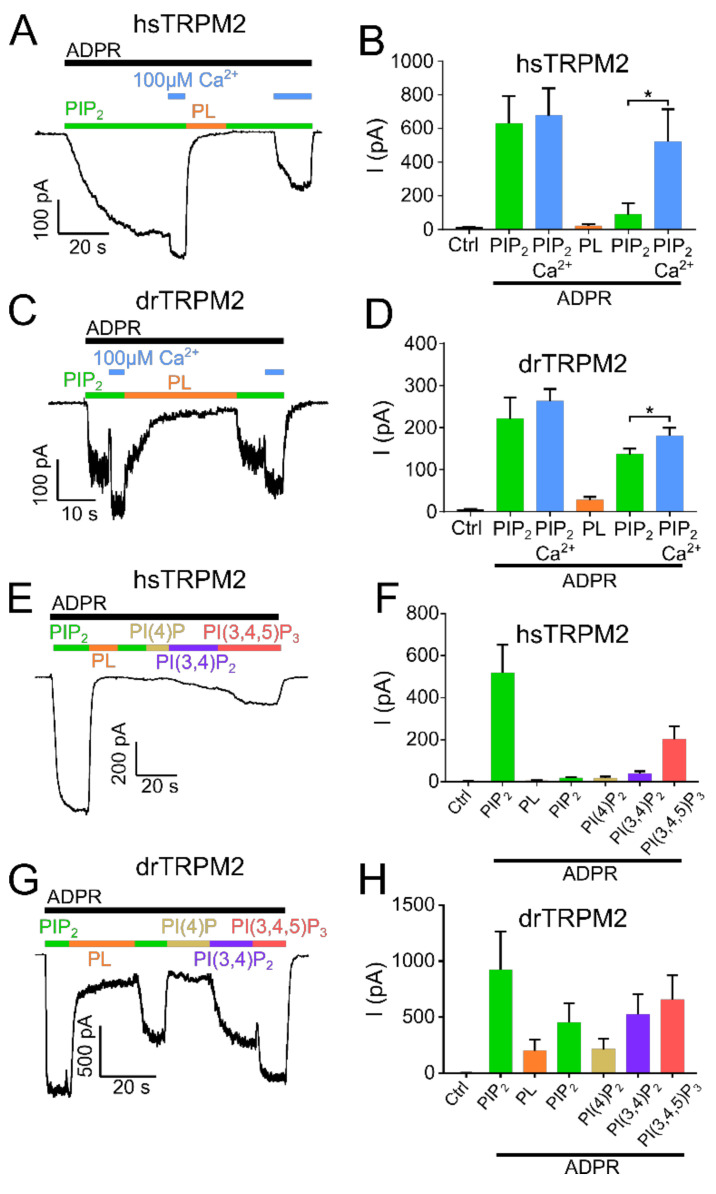
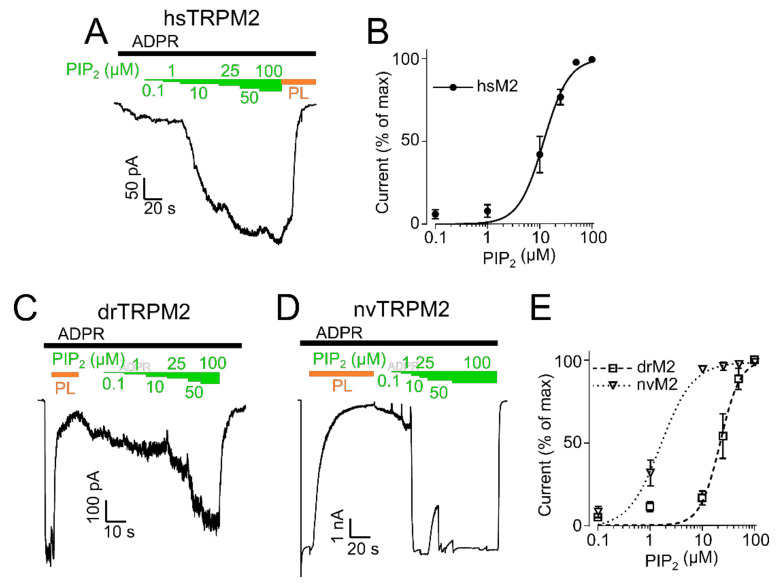
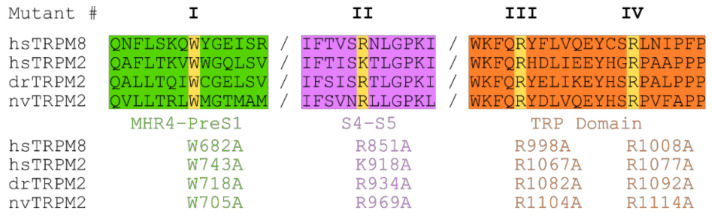
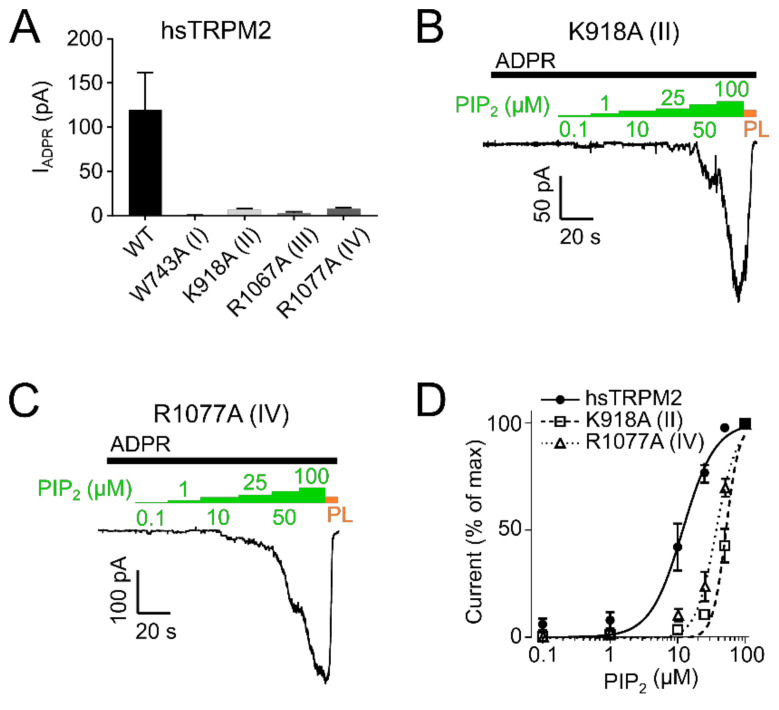
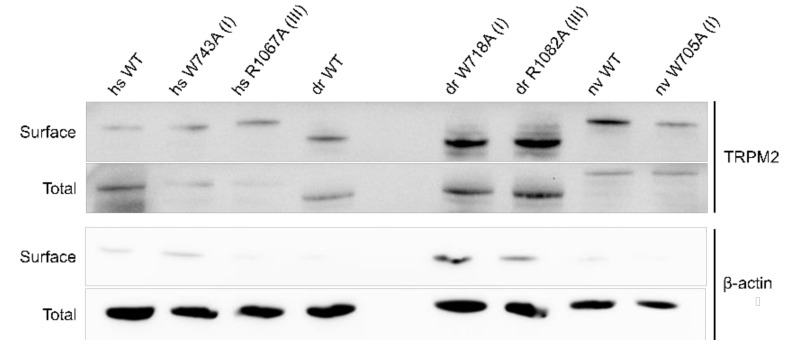
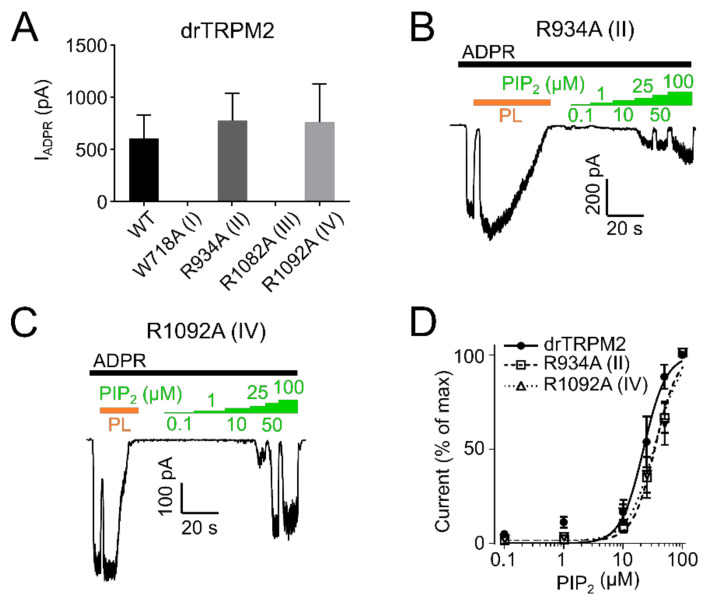
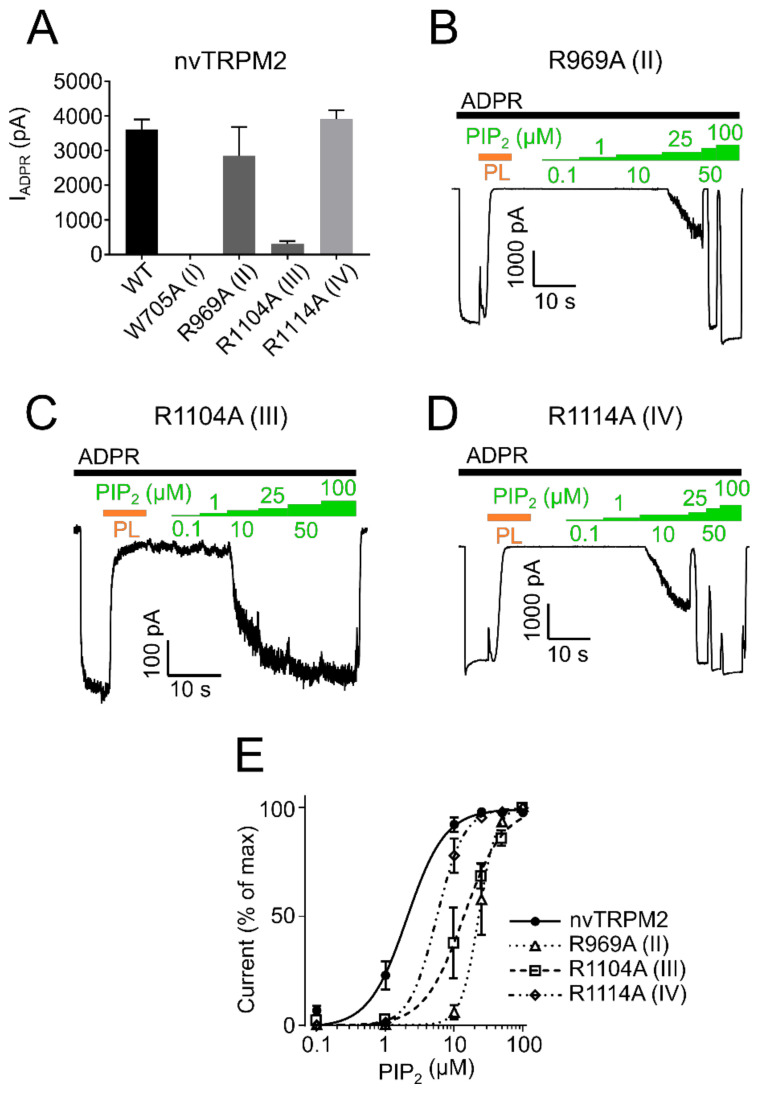
The human apoptosis channel TRPM2 is stimulated by intracellular ADR-ribose and calcium. Recent studies show pronounced species-specific activation mechanisms. Our aim was to analyse the functional effect of phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2), commonly referred to as PIP2, on different TRPM2 orthologues. Moreover, we wished to identify the interaction site between TRPM2 and PIP2. We demonstrate a crucial role of PIP2, in the activation of TRPM2 orthologues of man, zebrafish, and sea anemone. Utilizing inside-out patch clamp recordings of HEK-293 cells transfected with TRPM2, differential effects of PIP2 that were dependent on the species variant became apparent. While depletion of PIP2 via polylysine uniformly caused complete inactivation of TRPM2, restoration of channel activity by artificial PIP2 differed widely. Human TRPM2 was the least sensitive species variant, making it the most susceptible one for regulation by changes in intramembranous PIP2 content. Furthermore, mutations of highly conserved positively charged amino acid residues in the membrane interfacial cavity reduced the PIP2 sensitivity in all three TRPM2 orthologues to varying degrees. We conclude that the membrane interfacial cavity acts as a uniform PIP2 binding site of TRPM2, facilitating channel activation in the presence of ADPR and Ca2+ in a species-specific manner.
Keywords: PIP2; TRPM2; patch clamp; phosphoinositides; phospholipid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
