

doi: 10.1161/CIRCRESAHA.120.317943.
Epub 2021 Apr 30.
Nampt Potentiates Antioxidant Defense in Diabetic Cardiomyopathy
Affiliations
- PMID: 33928788
- PMCID: PMC8513534
- DOI: 10.1161/CIRCRESAHA.120.317943
Item in Clipboard
Nampt Potentiates Antioxidant Defense in Diabetic Cardiomyopathy
Circ Res.
.
Abstract
[Figure: see text].
Keywords: apoptosis; diabetic cardiomyopathies; fatty acids; fibrosis; oxidative stress.
Figures
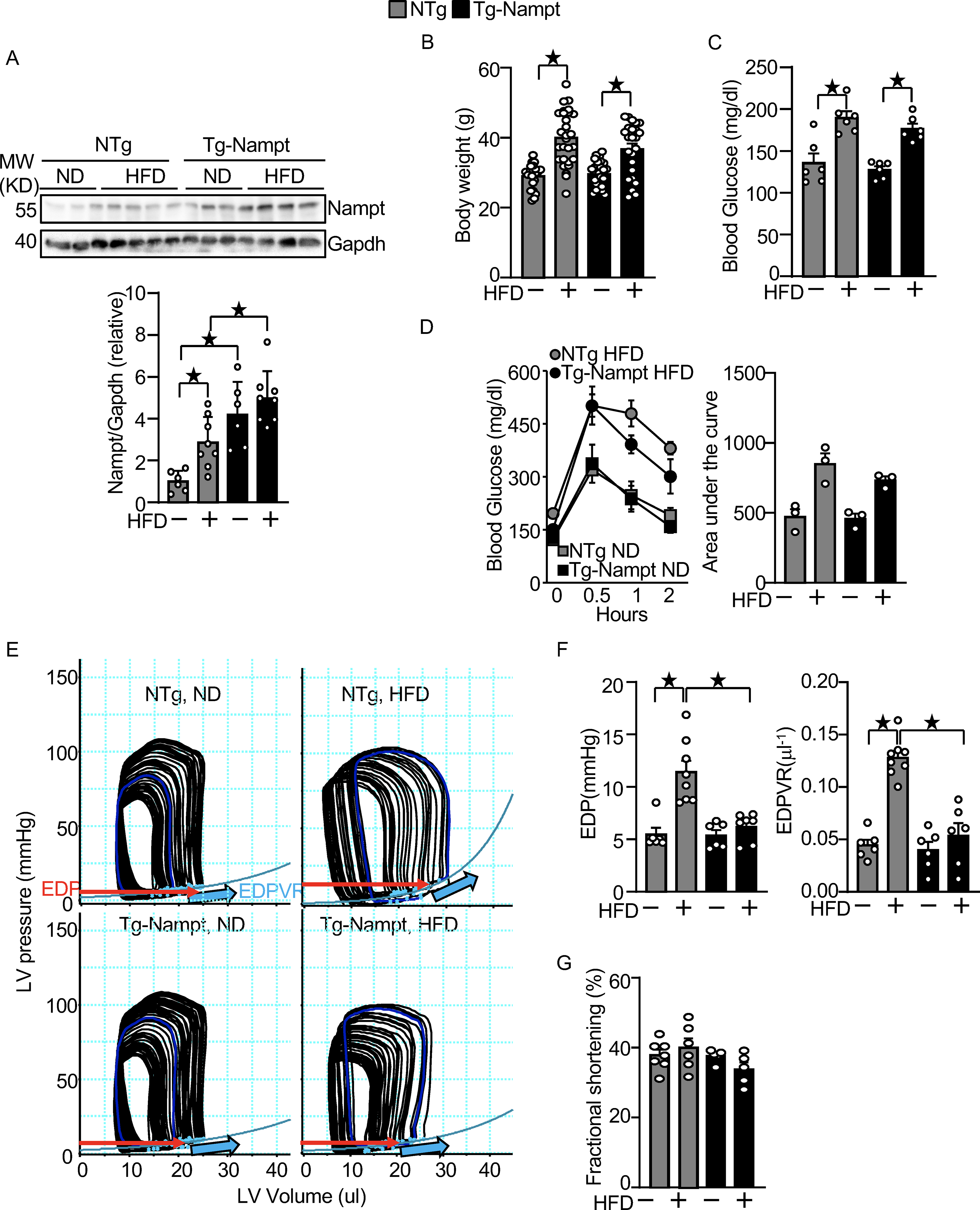
Nampt overexpression ameliorates HFD-induced diastolic dysfunction. (A) HFD consumption upregulates Nampt. Heart lysates were prepared from NTg and Tg-Nampt mice after 3 months of HFD consumption. Western blot analyses were performed with indicated antibodies. (B) Body weights in NTg and Tg-Nampt mice after 3 months of HFD consumption. The mice were fed with HFD beginning at 6–12 weeks of age for 3 months. (C) Blood glucose in NTg and Tg-Nampt mice after 3 months of HFD consumption. (D) Glucose tolerance in NTg and Tg-Nampt mice under ND feeding and 3 months of HFD consumption. (E) Representative PV loop results after 3 months of HFD consumption. (F) Increased EDP and EDPVR were observed after 3 months of HFD consumption in NTg but were normalized in Tg-Nampt mice. (E-F) Mice were anesthetized using pentobarbital (G) LV fractional shortening was preserved in Tg-Nampt mice fed ND and after 3 months of HFD consumption. Statistical significance was determined with ANOVA (A and F (EDPVR)), repeated measures ANOVA (B, C, D (Left) and G) and the Kruskal-Wallis test (D (right) and F (EDP)).
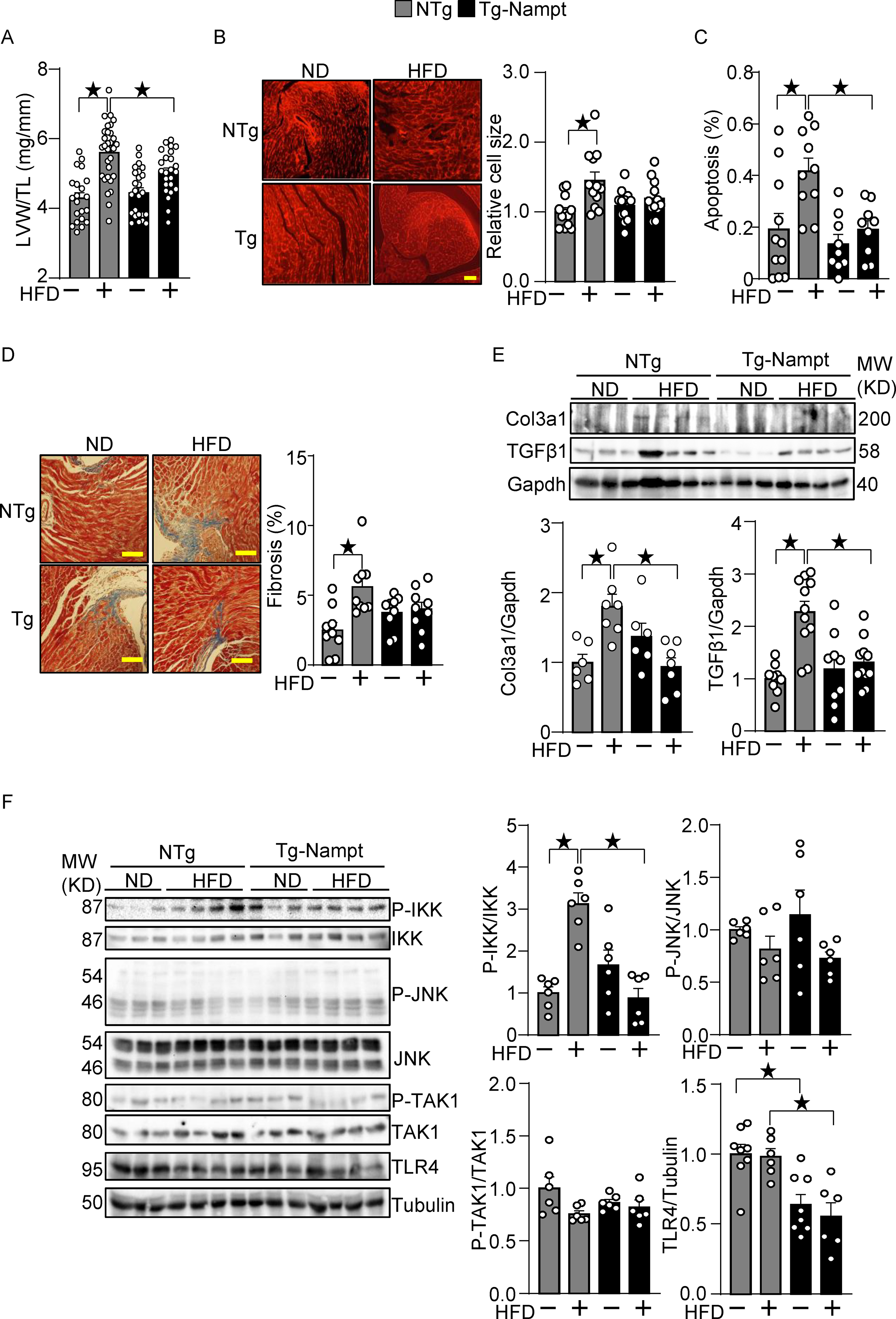
Nampt overexpression ameliorates HFD-induced cardiac pathologies relevant to diastolic dysfunction. (A-B) Nampt ameliorates HFD-induced left ventricular (LV) (A) and cardiomyocyte (B) hypertrophy. (A) LV hypertrophy indicated by LV weight (LVW)/tibial length (TL). (B) Relative cell size was evaluated, using wheat germ agglutinin (WGA) staining. (C) Nampt ameliorates HFD-induced apoptotic cell death. TUNEL-positive cells (apoptotic cells) were quantified as the number of TUNEL-positive/total number of nuclei (%). (D) Nampt ameliorates HFD-induced cardiac fibrosis. Cardiac fibrosis was evaluated using Masson’s trichrome staining. (Left) Representative images from each group are shown. Scale bars, 100 μm. (Right) Quantitative analysis of the fibrotic areas. (E) Nampt inhibits HFD-induced fibrosis markers. (F) Nampt inhibits HFD-induced IKK activation. Heart lysates were prepared from NTg and Tg-Nampt mice after 3 months of HFD feeding. Indicated pro-inflammatory signal regulators were examined with Western blot analyses. Statistical significance was determined with ANOVA (A, B, C, E and F (P-JNK/JNK, P-TAK1/TAK1 and TLR4/Tubulin)) and the Kruskal-Wallis test (D, F (P-IKK/IKK)).
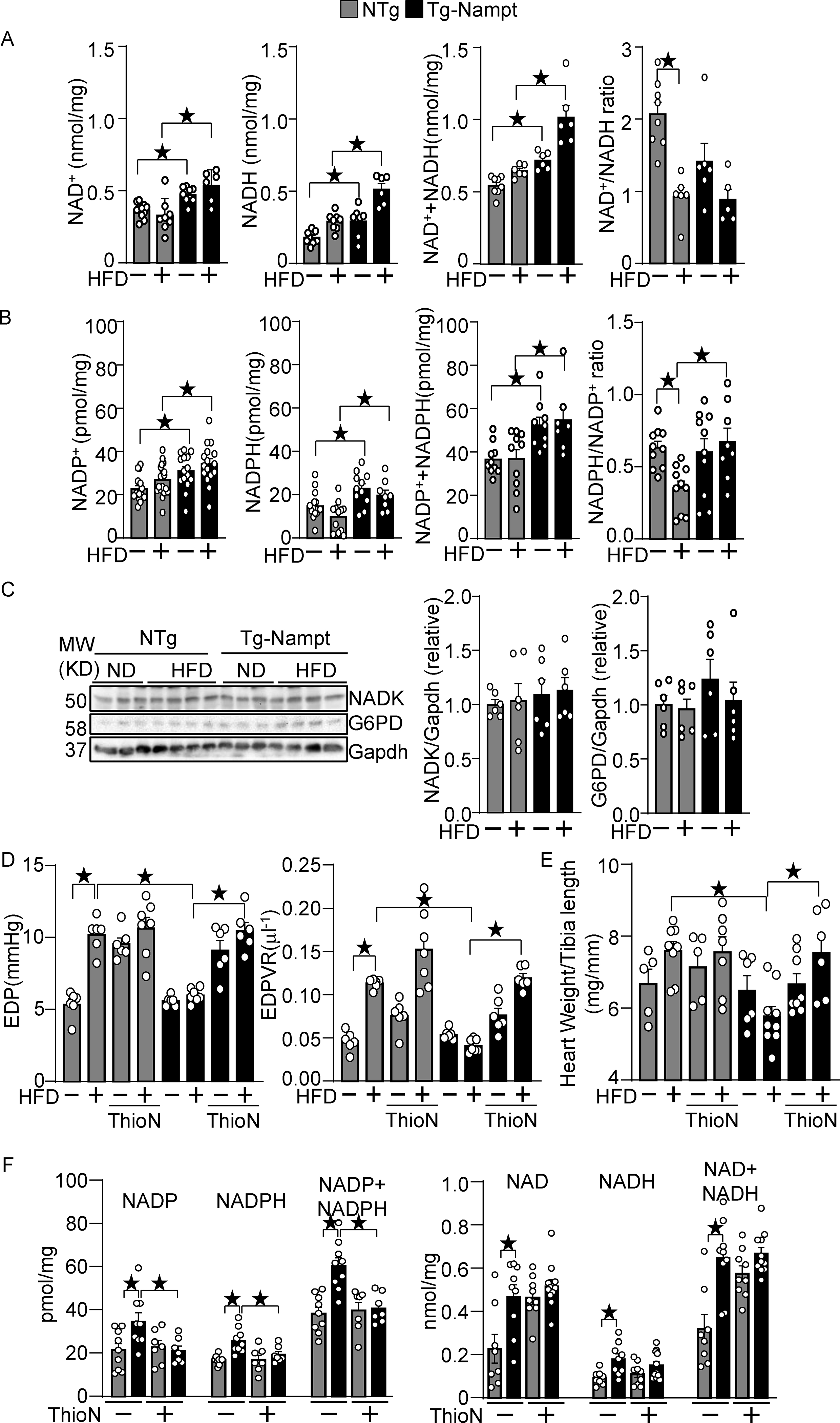
Nampt overexpression upregulates NAD(H) and NADP(H). (A) The levels of NAD+, NADH and total NAD++NADH were examined in Tg-Nampt mice after either normal diet or HFD consumption. (B) The levels of NADP+, NADPH, and total NADP++NADPH and the NADPH/NADP+ ratio were examined in Tg-Nampt mice under HFD feeding conditions. (C) Neither Nampt overexpression nor HFD significantly affects the level of NADK or G6PD (D) Nampt overexpression attenuates HFD-induced diastolic dysfunction in an NADK-dependent manner. Mice were anesthetized with Avertin. (E) Nampt overexpression attenuates HFD-induced cardiac hypertrophy in an NADK-dependent manner. (F) ThioN, an NADK inhibitor, inhibits Nampt-induced NADP(H), but not NAD(H) production. Statistical significance was determined with ANOVA (A-F).
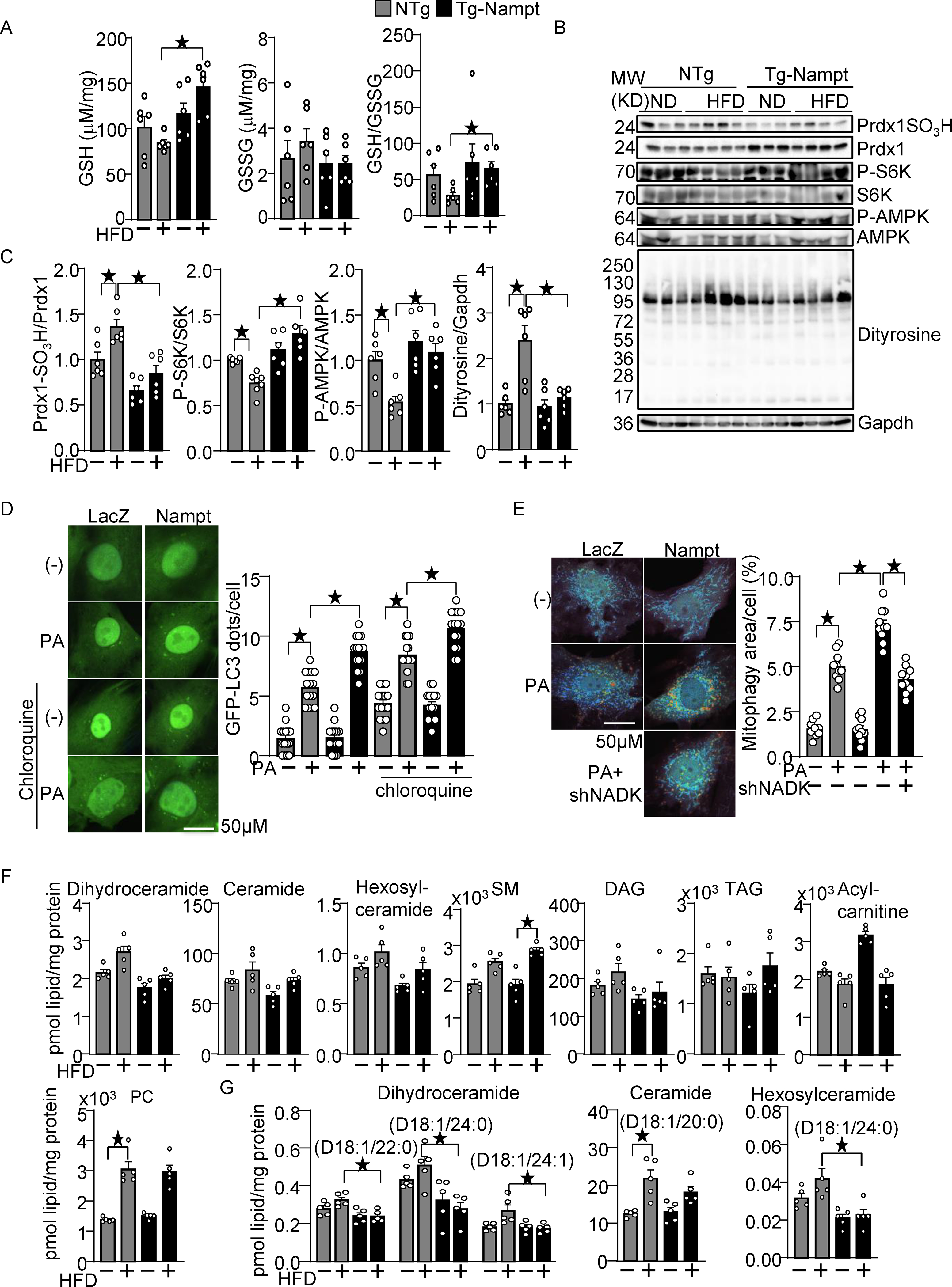
Nampt overexpression attenuates HFD-induced oxidative stress. (A) Nampt overexpression attenuates HFD-induced GSH oxidation. The levels of GSH and GSSG and the GSH/GSSG ratio were examined in Tg-Nampt mice after 3 months of HFD consumption. (B-C) Nampt overexpression attenuates HFD-induced inhibition of Trx1 substrates. Heart lysates were prepared from NTg and Tg-Nampt mice after 3 months of HFD consumption. Western blot analyses were performed with indicated antibodies. (D) Nampt promotes palmitic acid (PA)-induced autophagy. After 2 days of adenovirus transduction of Nampt and GFP-LC3, cardiomyocytes were treated with 100 μM PA and 10 μM chloroquine for 4 hours. GFP-LC3 dots per cell were counted. (E) Nampt promotes PA-induced mitophagy in an NADK-dependent manner. After 3 days of adenovirus transduction of Nampt and Mito-Keima, NADK was knocked down with shNADK. After 3 days, the cardiomyocytes were treated with 100 μmol/L PA for 24 hours. Areas with high 561/457 nm ratios, indicating mitophagy, were measured. (F-G) The effect of Nampt overexpression upon myocardial contents of various lipid species in the presence or absence of HFD consumption. (F) The effect upon the total amount of indicated lipid species. (G) The effect upon several specific subspecies of dihydroceramide, ceramide and hexosylceramide. Statistical significance was determined with ANOVA (A (GSH and GSSG), and C-E) and the Kruskal-Wallis test (A (GSH/GSSG), F and G.
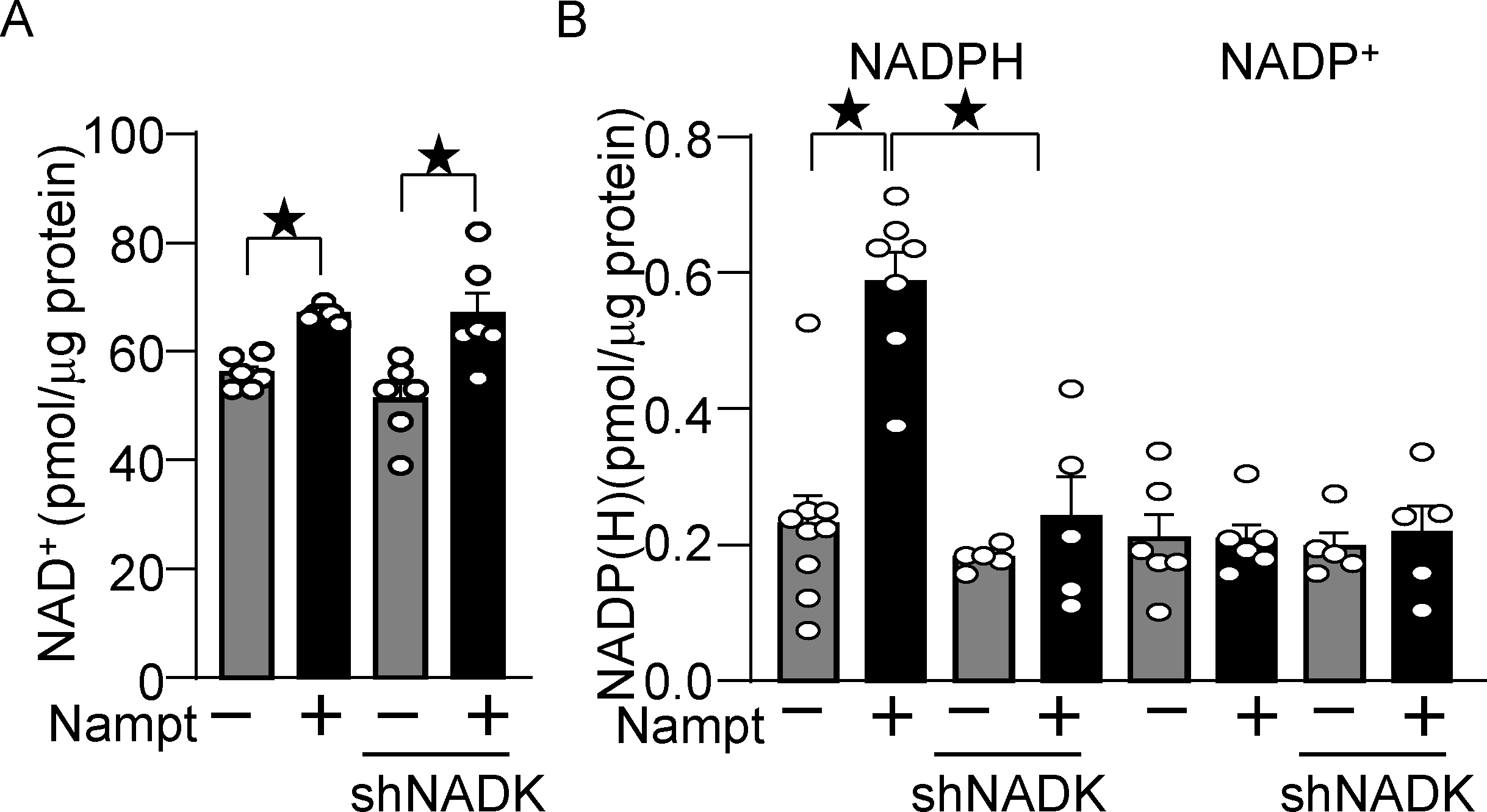
Nampt overexpression upregulates NADP(H) in an NADK-dependent manner. (A) Nampt upregulates NAD+, which is not affected by NADK knockdown. Statistical significance was determined with ANOVA. (B) Nampt upregulates NADPH in an NADK-dependent manner. After 1 day of adenovirus vector (Nampt) transduction, shNADK was transfected into cardiomyocytes. After 2 days of adenovirus transduction, the levels of NAD+, NADP+ and NADPH were examined. Statistical significance was determined with ANOVA (A and B (NADP)) and the Kruskal-Wallis test (B (NADPH)).
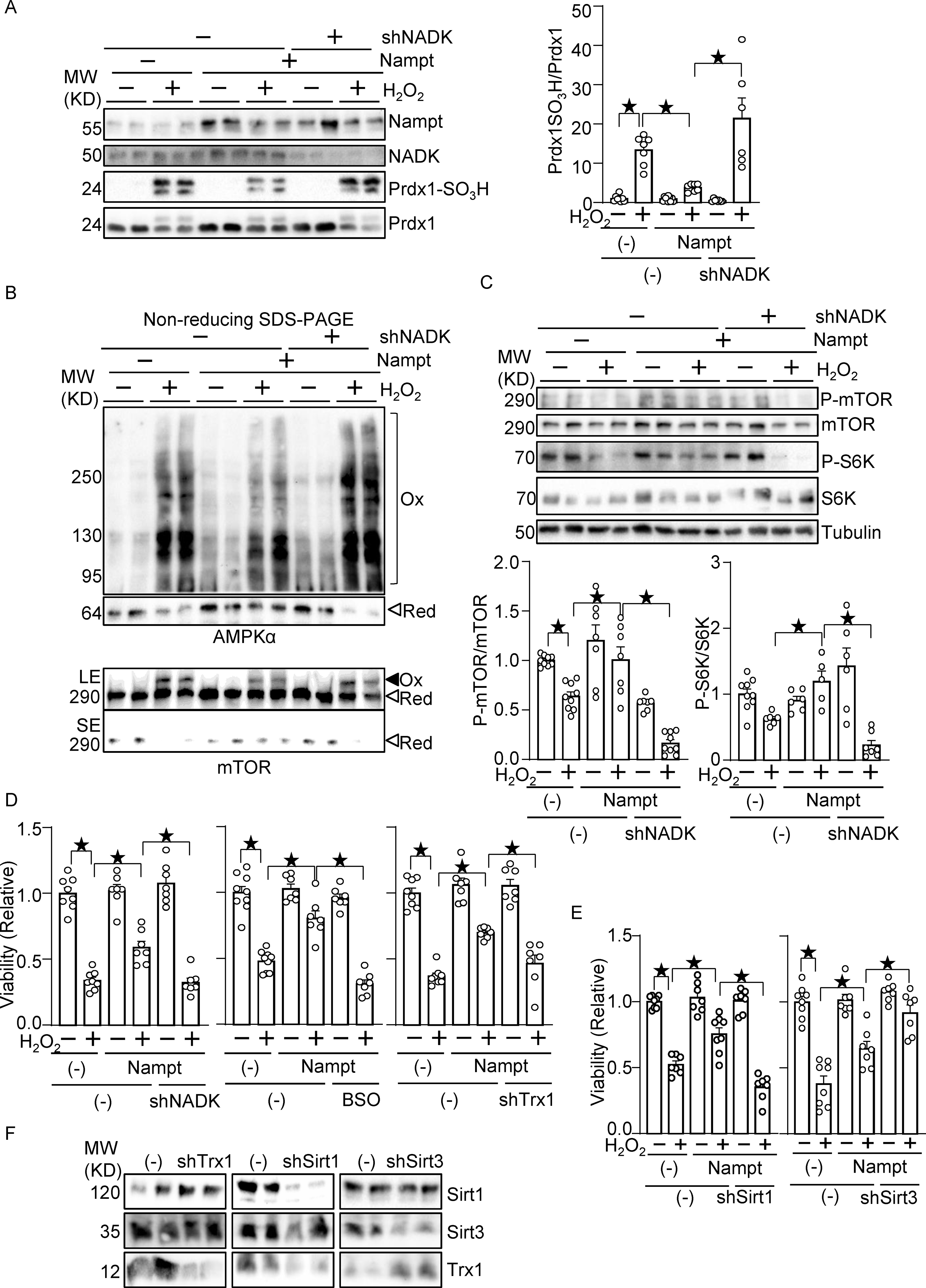
Nampt alleviates oxidative stress in an NADK-dependent manner. (A) Nampt prevents H2O2-induced Prdx1 oxidation in an NADK-dependent manner. (B) Nampt prevents H2O2-induced intermolecular disulfide bond formation in AMPKα and mTOR in an NADK-dependent manner (Ox: oxidized, Red: reduced, LE: long exposure, SE: short exposure). (C) Nampt prevents H2O2-induced mTOR inhibition in an NADK-dependent manner. After 1 day of adenovirus vector (Nampt) transduction, shNADK was transfected into cardiomyocytes. After 1 or 2 days of transfection, cells were treated with 100 μM H2O2 for 30 minutes. Western blot analyses were performed following SDS-PAGE under reducing (A and C) and non-reducing (B) conditions using indicated antibodies. (D) Nampt prevents H2O2-induced myocyte cell death, an effect that is partly abolished by inhibition of NADK, GSH synthesis or Trx1. (Left) After adenovirus transduction and shRNA transfection as described above, cells were treated with 100 μmol/L H2O2 for 6 hours. Cell viability was assessed by trypan blue dye exclusion. (Middle) Cells were treated with buthionine sulfoximine (BSO, 100 μmol/L), a GSH synthesis inhibitor, for 16 hours prior to H2O2 treatment. (Right) Adenovirus vector harboring shTrx1 was transduced together with that for Nampt. (E) Sirt1, but not Sirt3, mediates the protective effects of Nampt against H2O2-induced myocyte cell death. After adenovirus transduction and shRNA transfection, cells were treated with 100 μmol/L H2O2 for 6 hours. Cell viability was assessed by trypan blue dye exclusion. (F) Western blot showing knockdown of Trx1, Sirt1 and Sirt3. Statistical significance was determined with ANOVA (A-E).
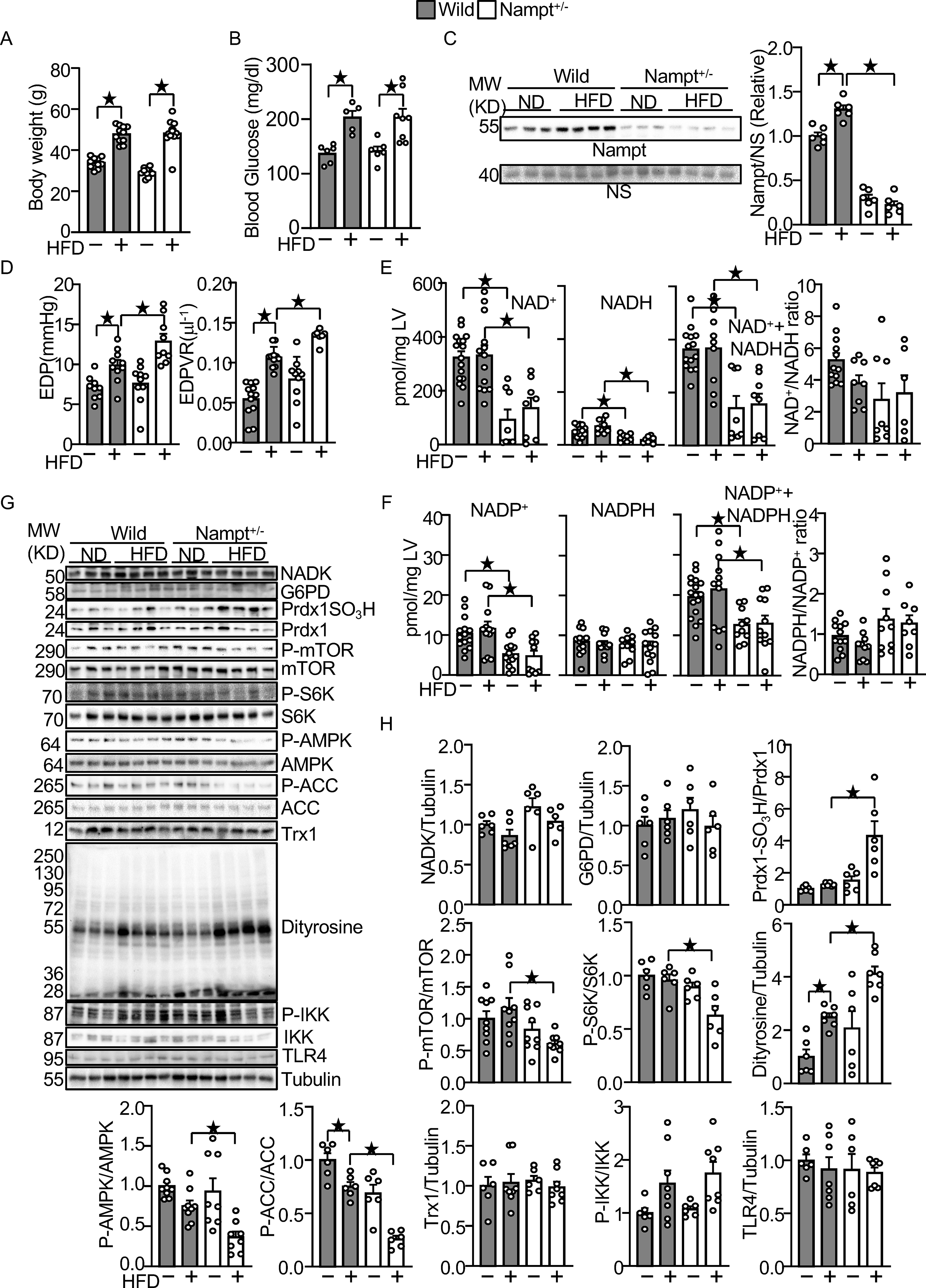
Endogenous Nampt prevents HFD-induced diastolic dysfunction and oxidative stress. (A) Body weights in wild type and Nampt+/− mice under HFD feeding conditions. (B) Blood glucose in wild type and Nampt+/− mice under HFD feeding conditions (C) HFD-induced Nampt expression is inhibited in Nampt+/− mice. NS: Non-specific band. (D) HFD-induced diastolic dysfunction is exacerbated in Nampt+/− mice. (E) Endogenous Nampt maintains NAD(H). (F) Endogenous Nampt maintains NADP. (G) Neither Nampt knockdown nor HFD significantly affects the level of NADK and G6PD. (H) Nampt knockdown promotes HFD-induced inhibition of Trx1 substrates. Heart lysates were prepared from WT and Nampt+/− mice after 3 months of HFD consumption. Western blot analyses were performed with indicated antibodies. Statistical significance was determined with ANOVA (C, E (NADH), F (NADPH, NADP+NADPH, NADPH/NADP), G, H (NADK/Tubulin, G6PD/Tubulin, PrdxSO3H/Prdx1, P-mTOR/mTOR. P-S6K/S6K, P-AMPKα/AMPKα, P-ACC/ACC, Dityrosine/Tubulin and P-IKK/IKK), repeated measures ANOVA (A, B) and the Kruskal-Wallis test (D, E (NAD, NAD+NADH, NAD/NADH), F (NADP) and H (Trx1/Tubulin and TLR4/Tubulin).
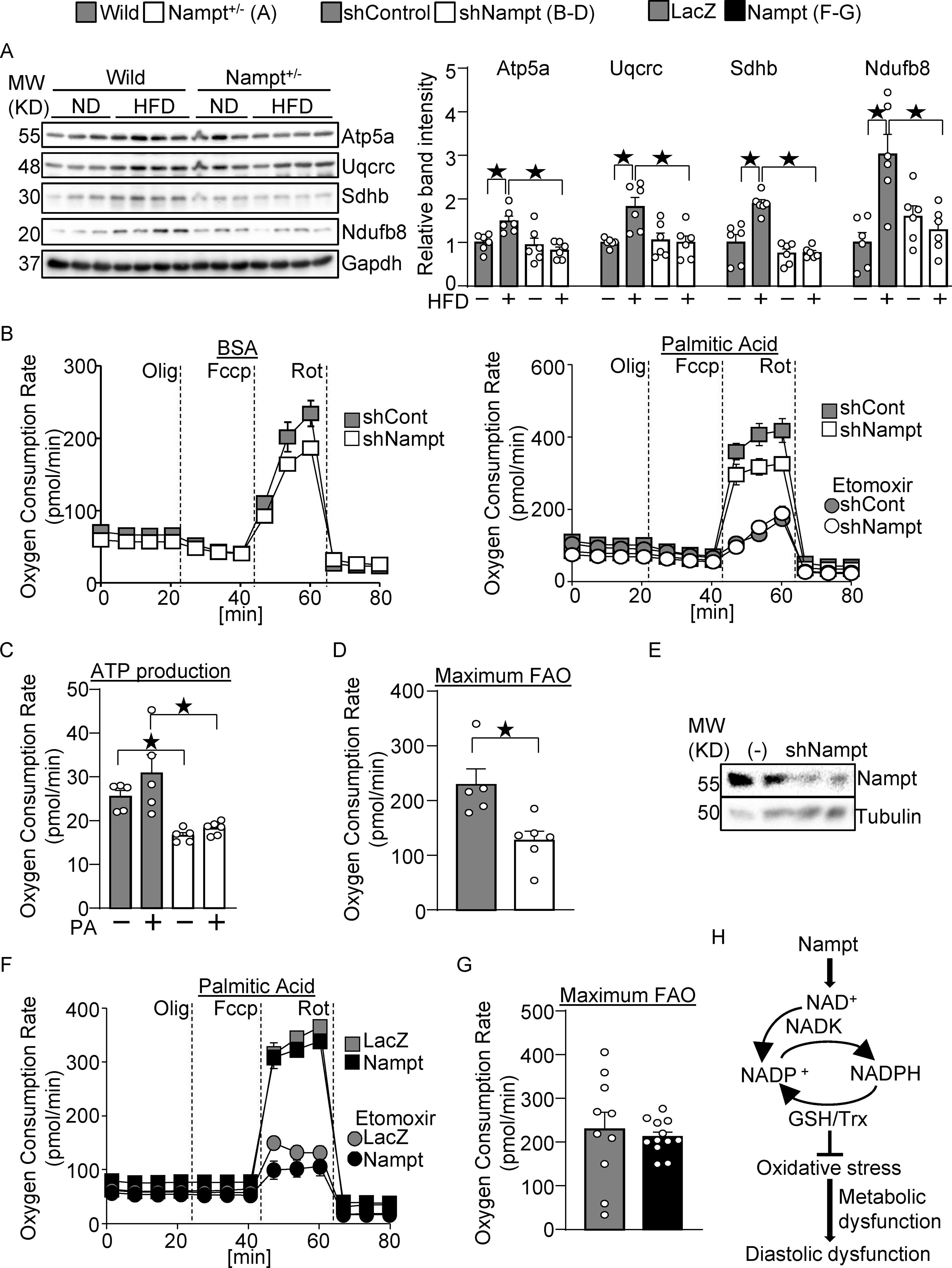
Nampt potentiates metabolic ability. (A) Nampt knockdown suppresses mitochondrial proteins during HFD consumption. Heart lysates were prepared from WT and Nampt+/− mice after 3 months of HFD consumption. Western blot analyses were performed with indicated antibodies. (B-D) Nampt potentiates ATP production and fatty acid oxidation. Cardiomyocytes were transduced with shNampt. Oxygen consumption rate was examined in cardiomyocytes incubated with 100 μmol/L palmitic acid (PA). ATP production coupled oxygen consumption (C) and maximum fatty acid oxidation capacity (D) are shown. (C-D) N=5–6. (E) Western blot showing knockdown of Nampt by adenovirus vector of shNampt. (F-G) Cardiomyocytes were transduced with Ad-Nampt. Oxygen consumption rate was examined in cardiomyocytes incubated with 100 μmol/L palmitic acid (PA). Statistical significance was determined with ANOVA (A), the Kruskal-Wallis test (C), the Mann-Whitney U test (G), and the Student’s t test (G). (H) A schematic representation of the current hypothesis. Nampt promotes NAD production, which promotes NADP production via NADK. Increased NADP(H) confers resistance against oxidative stress, possibly through the GSH and Trx systems, which may in turn attenuate diastolic dysfunction.
References
-
- Engin A The Definition and Prevalence of Obesity and Metabolic Syndrome. Adv Exp Med Biol. 2017;960:1–17. - PubMed
-
- Paolillo S, Marsico F, Prastaro M, Renga F, Esposito L, De Martino F, Di Napoli P, Esposito I, Ambrosio A, Ianniruberto M, Mennella R, Paolillo R and Gargiulo P. Diabetic Cardiomyopathy: Definition, Diagnosis, and Therapeutic Implications. Heart Fail Clin. 2019;15:341–347. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
