

Promotion of cholangiocarcinoma growth by diverse cancer-associated fibroblast subpopulations
- PMID: 33930309
- PMCID: PMC8241235
- DOI: 10.1016/j.ccell.2021.03.012
Promotion of cholangiocarcinoma growth by diverse cancer-associated fibroblast subpopulations
Erratum in
-
Promotion of cholangiocarcinoma growth by diverse cancer-associated fibroblast subpopulations.Cancer Cell. 2021 Jun 14;39(6):883. doi: 10.1016/j.ccell.2021.05.010. Cancer Cell. 2021. PMID: 34129825 Free PMC article. No abstract available.
Abstract
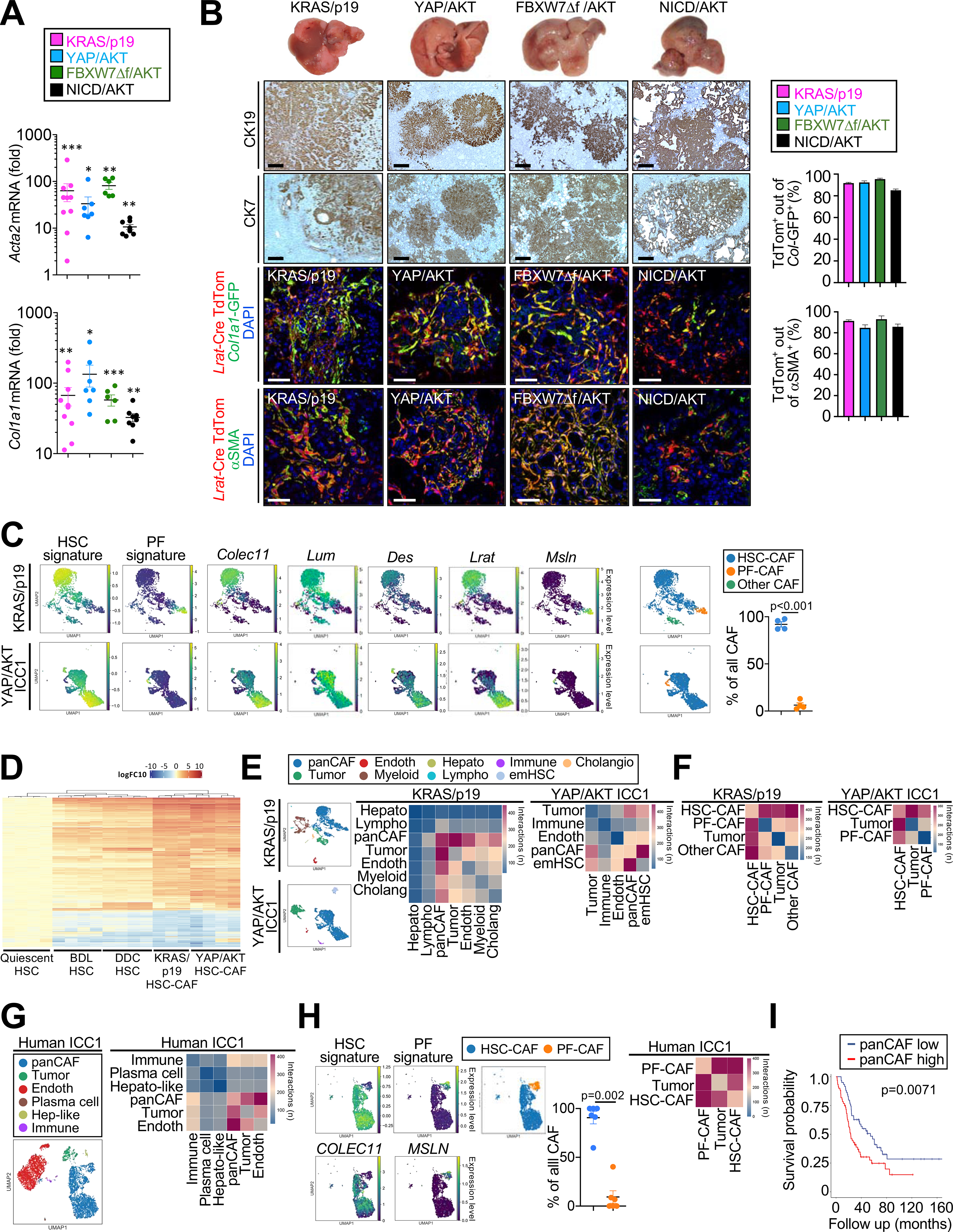
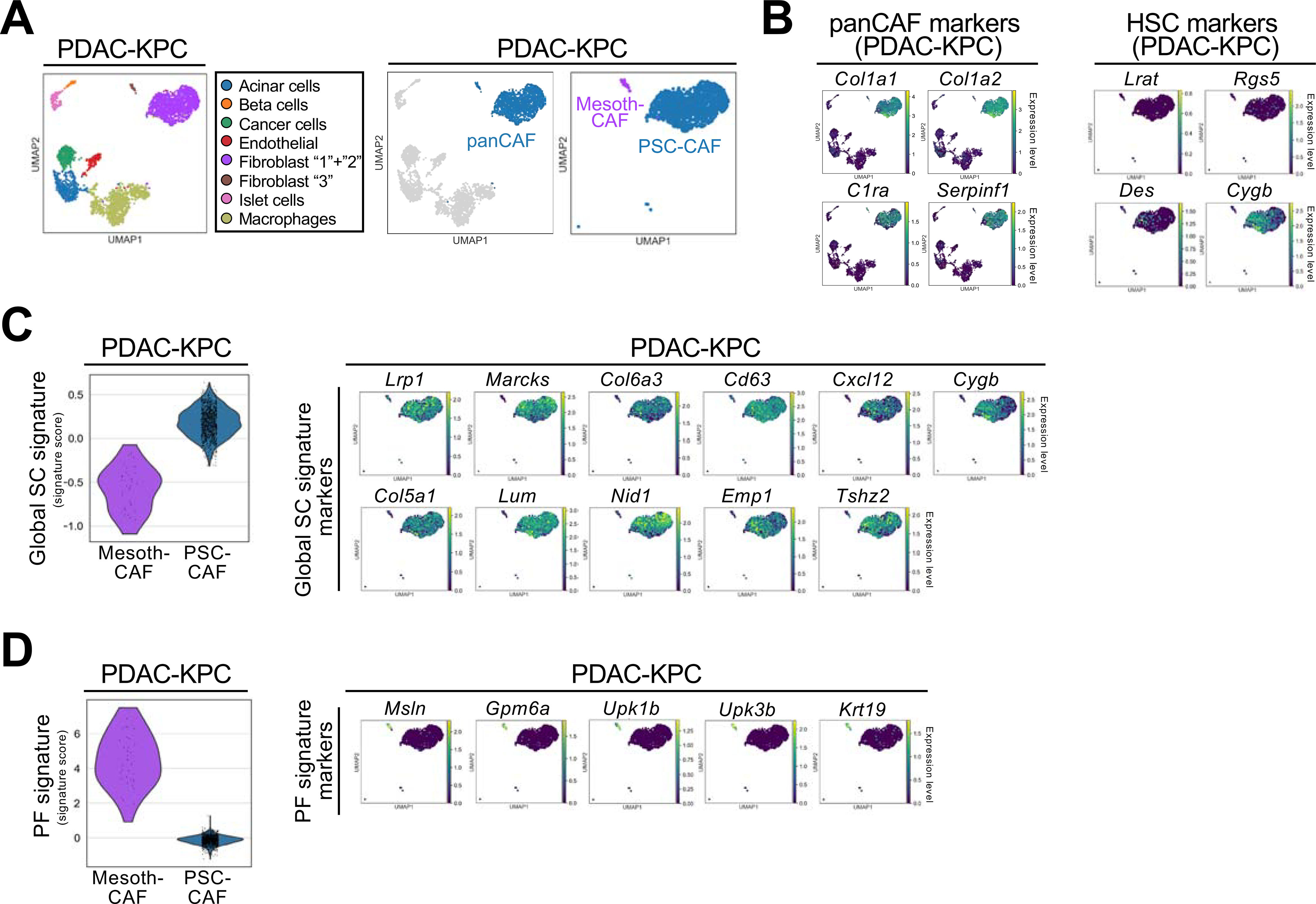
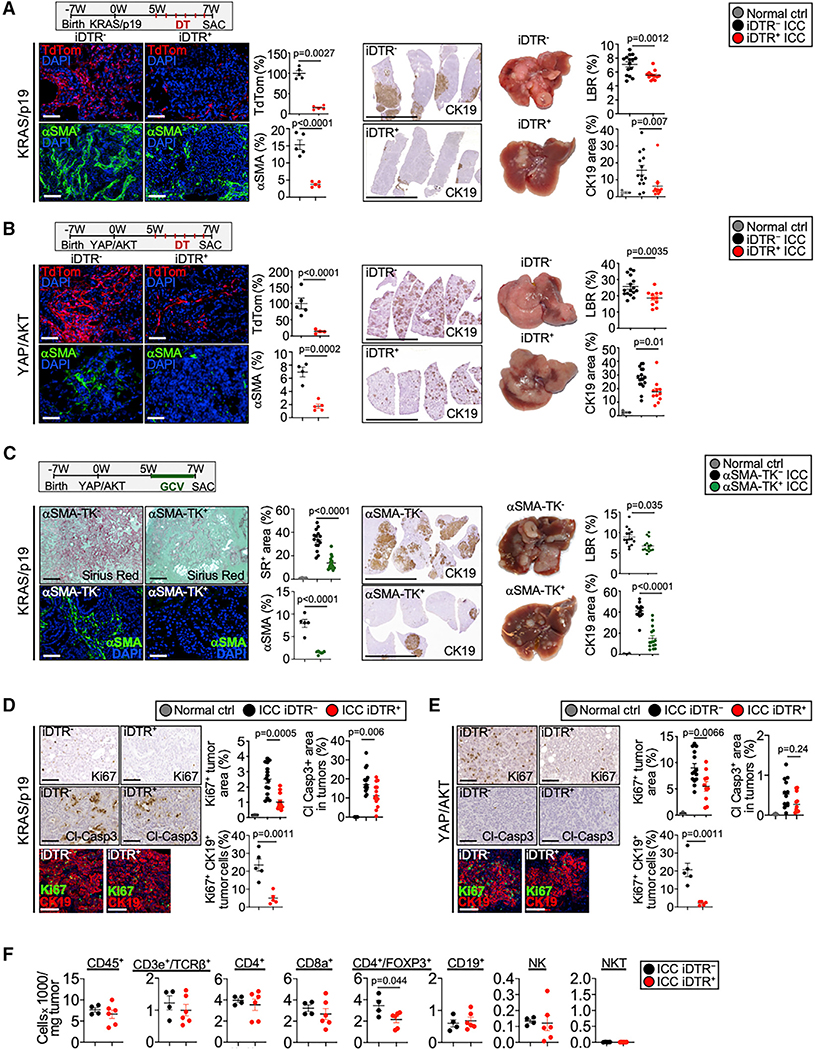
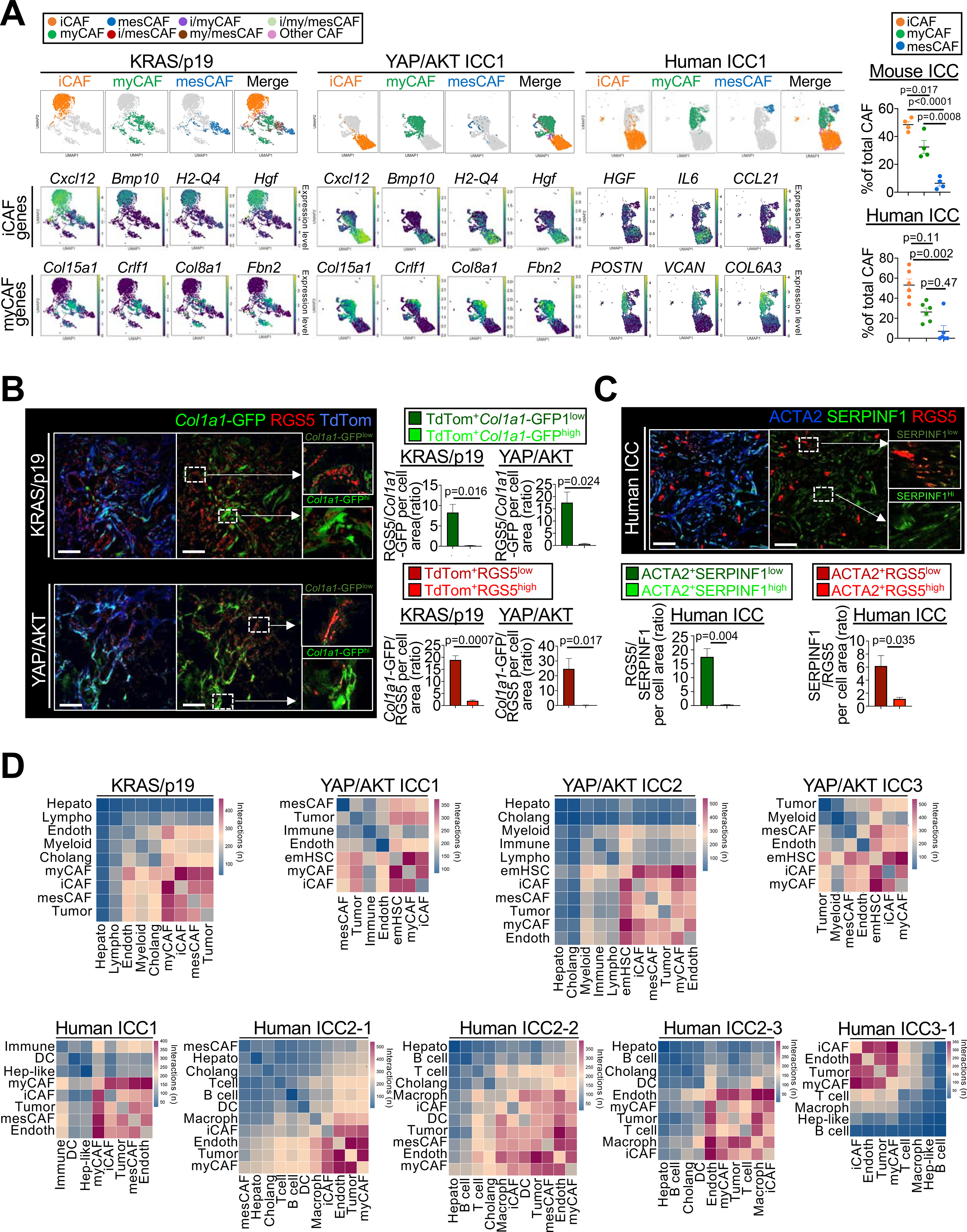
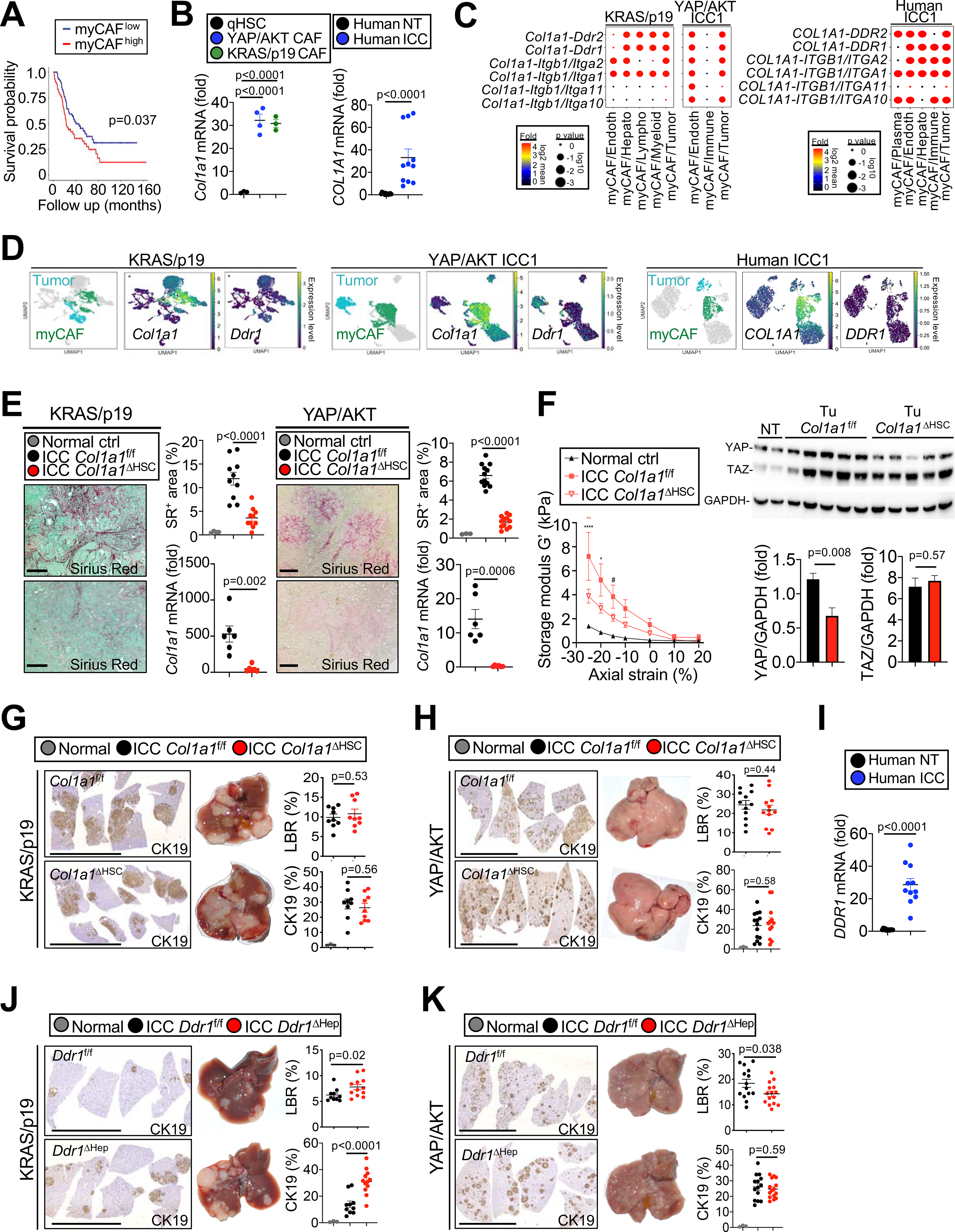
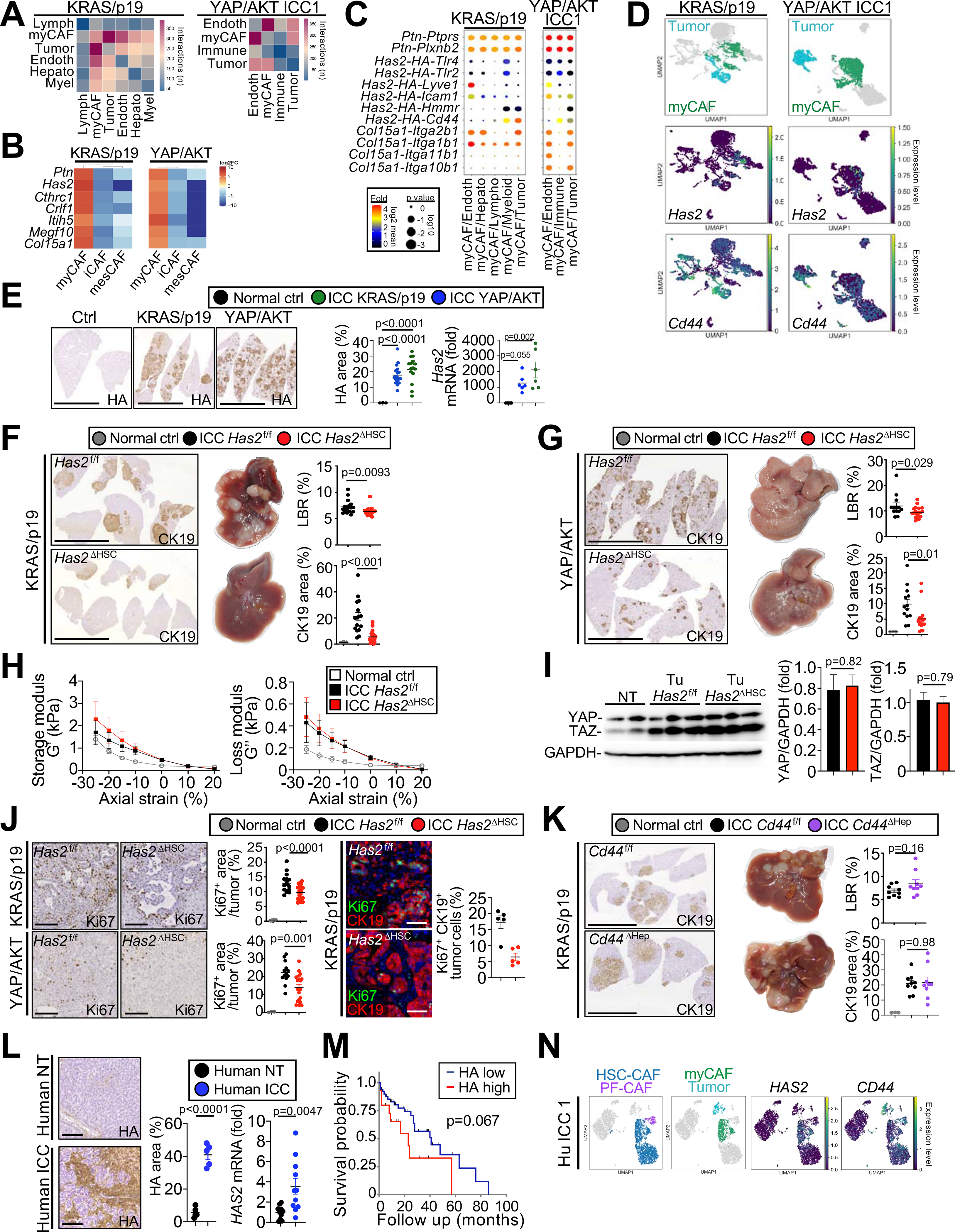
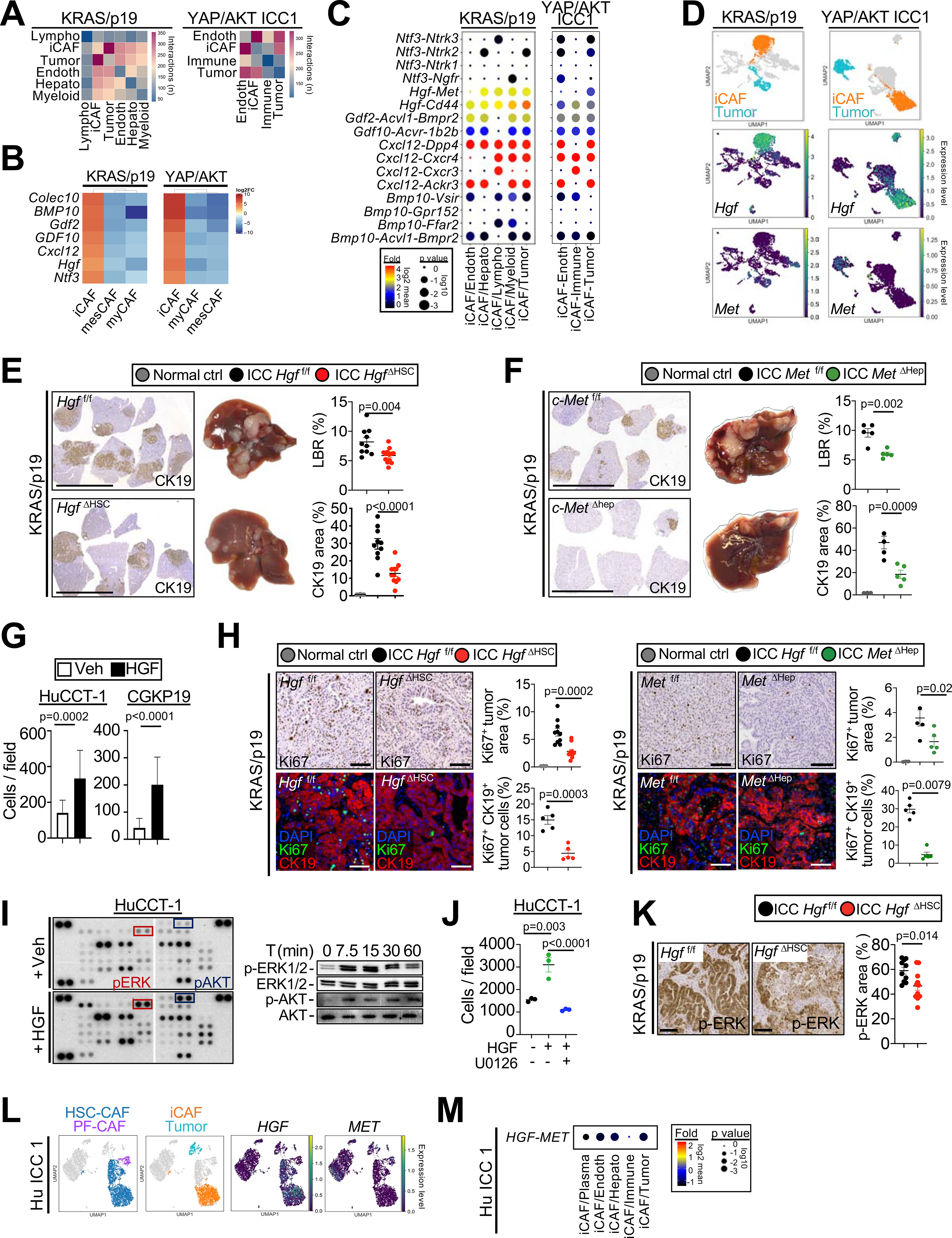
Cancer-associated fibroblasts (CAF) are a poorly characterized cell population in the context of liver cancer. Our study investigates CAF functions in intrahepatic cholangiocarcinoma (ICC), a highly desmoplastic liver tumor. Genetic tracing, single-cell RNA sequencing, and ligand-receptor analyses uncovered hepatic stellate cells (HSC) as the main source of CAF and HSC-derived CAF as the dominant population interacting with tumor cells. In mice, CAF promotes ICC progression, as revealed by HSC-selective CAF depletion. In patients, a high panCAF signature is associated with decreased survival and increased recurrence. Single-cell RNA sequencing segregates CAF into inflammatory and growth factor-enriched (iCAF) and myofibroblastic (myCAF) subpopulations, displaying distinct ligand-receptor interactions. myCAF-expressed hyaluronan synthase 2, but not type I collagen, promotes ICC. iCAF-expressed hepatocyte growth factor enhances ICC growth via tumor-expressed MET, thus directly linking CAF to tumor cells. In summary, our data demonstrate promotion of desmoplastic ICC growth by therapeutically targetable CAF subtype-specific mediators, but not by type I collagen.
Keywords: CellPhoneDB; HGF; KRAS; YAP; cholangiocarcinoma; immune; mechanosensitive; single cell; stiffness; tumor microenvironment.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.C. is founder, equity holder, and consultant of DarwinHealth. Columbia University is an equity holder in DarwinHealth.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R35 CA209896/CA/NCI NIH HHS/United States
- P30 DK026743/DK/NIDDK NIH HHS/United States
- P01 CA117969/CA/NCI NIH HHS/United States
- R01 DK085252/DK/NIDDK NIH HHS/United States
- U54 CA193417/CA/NCI NIH HHS/United States
- S10 OD012351/OD/NIH HHS/United States
- S10 OD021764/OD/NIH HHS/United States
- R01 CA228483/CA/NCI NIH HHS/United States
- R35 CA197745/CA/NCI NIH HHS/United States
- R01 CA190606/CA/NCI NIH HHS/United States
- R03 DK101863/DK/NIDDK NIH HHS/United States
- P30 ES013508/ES/NIEHS NIH HHS/United States
- P01 CA087497/CA/NCI NIH HHS/United States
- R01 CA190844/CA/NCI NIH HHS/United States
- R61 NS109407/NS/NINDS NIH HHS/United States
- P30 CA013696/CA/NCI NIH HHS/United States
- P01 CA233452/CA/NCI NIH HHS/United States
- U01 CA217858/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
