

LanCLs add glutathione to dehydroamino acids generated at phosphorylated sites in the proteome
- PMID: 33932340
- PMCID: PMC8209957
- DOI: 10.1016/j.cell.2021.04.001
LanCLs add glutathione to dehydroamino acids generated at phosphorylated sites in the proteome
Abstract
Enzyme-mediated damage repair or mitigation, while common for nucleic acids, is rare for proteins. Examples of protein damage are elimination of phosphorylated Ser/Thr to dehydroalanine/dehydrobutyrine (Dha/Dhb) in pathogenesis and aging. Bacterial LanC enzymes use Dha/Dhb to form carbon-sulfur linkages in antimicrobial peptides, but the functions of eukaryotic LanC-like (LanCL) counterparts are unknown. We show that LanCLs catalyze the addition of glutathione to Dha/Dhb in proteins, driving irreversible C-glutathionylation. Chemo-enzymatic methods were developed to site-selectively incorporate Dha/Dhb at phospho-regulated sites in kinases. In human MAPK-MEK1, such "elimination damage" generated aberrantly activated kinases, which were deactivated by LanCL-mediated C-glutathionylation. Surveys of endogenous proteins bearing damage from elimination (the eliminylome) also suggest it is a source of electrophilic reactivity. LanCLs thus remove these reactive electrophiles and their potentially dysregulatory effects from the proteome. As knockout of LanCL in mice can result in premature death, repair of this kind of protein damage appears important physiologically.
Keywords: C-glutathionylation; LanCL; MEK1; dehydroalanine; dehydrobutyrine; eliminylome; lanthionine; phosphoThr lyase; protein damage.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
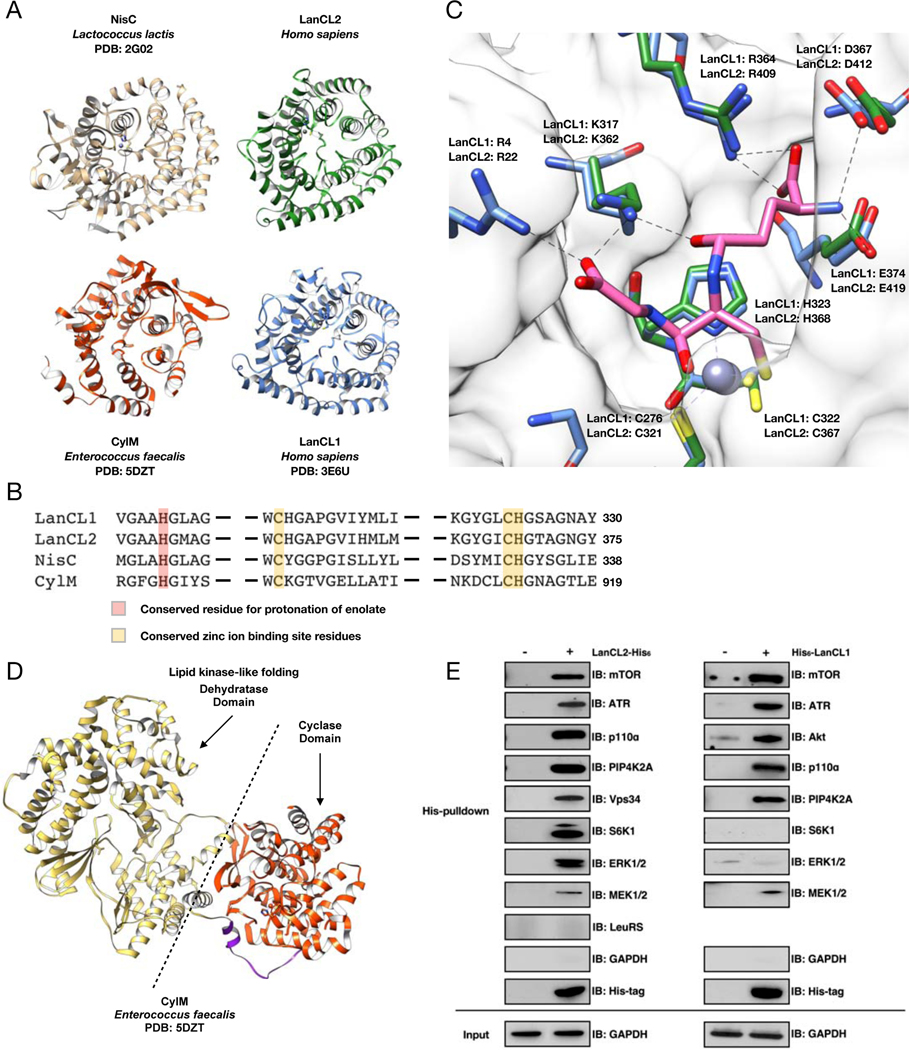
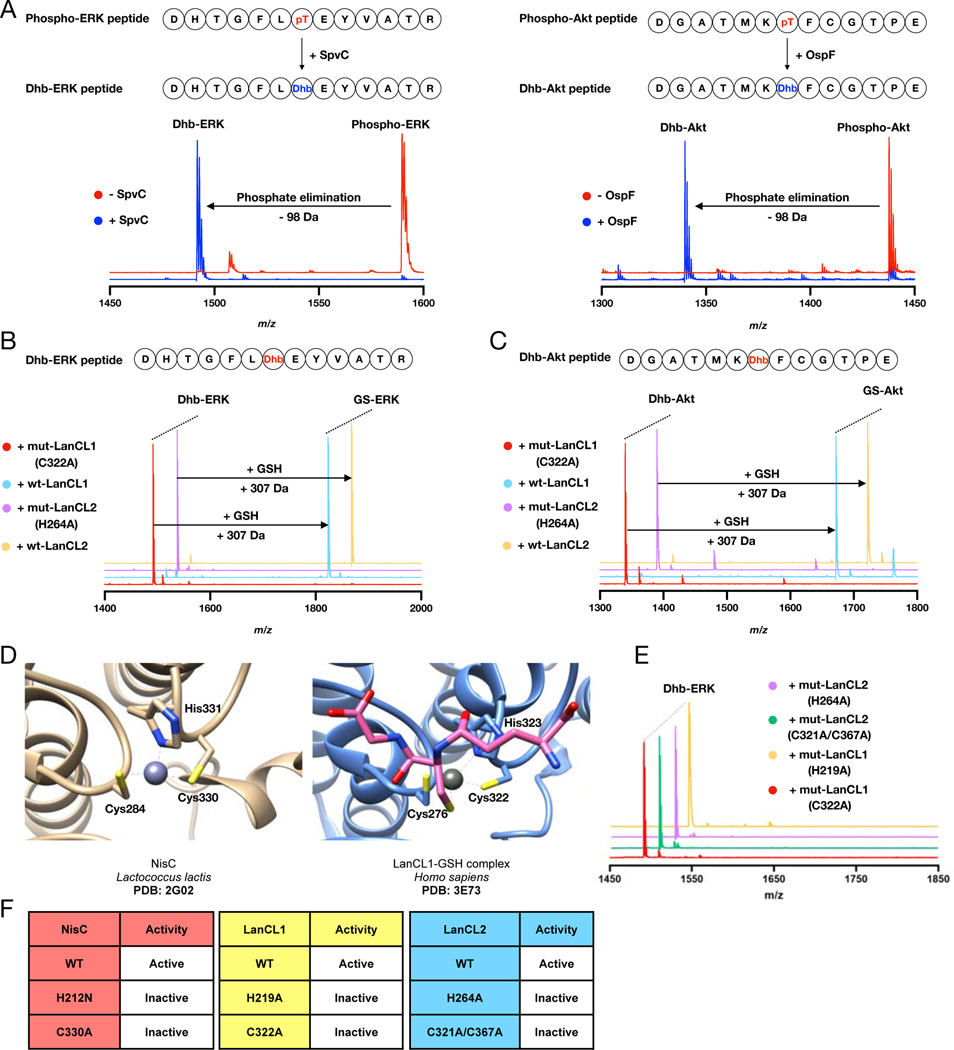
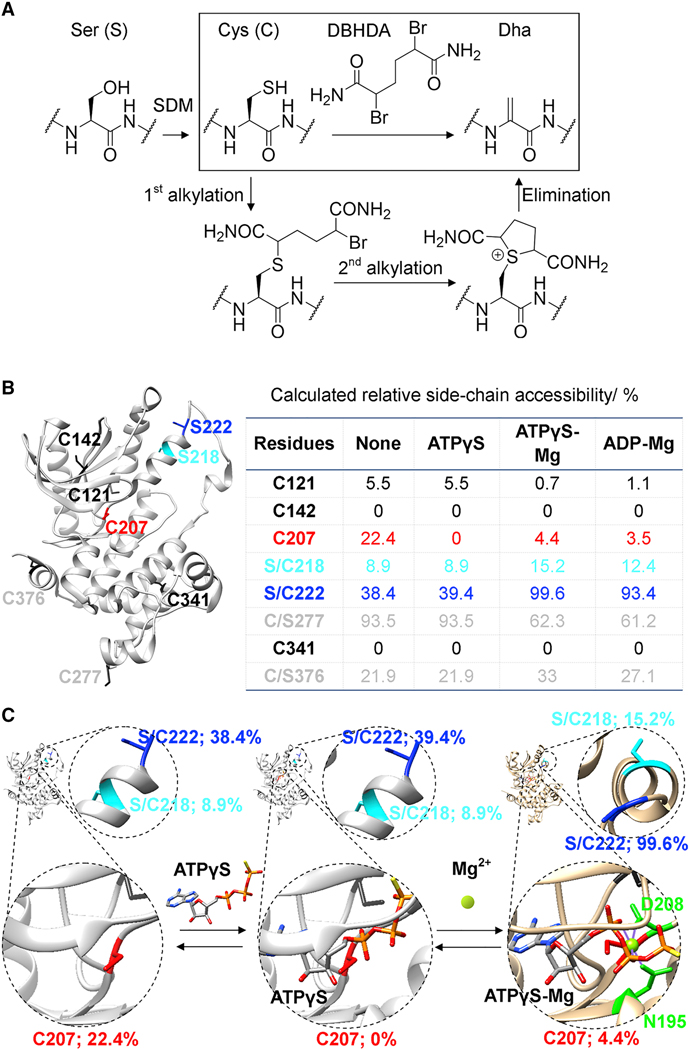
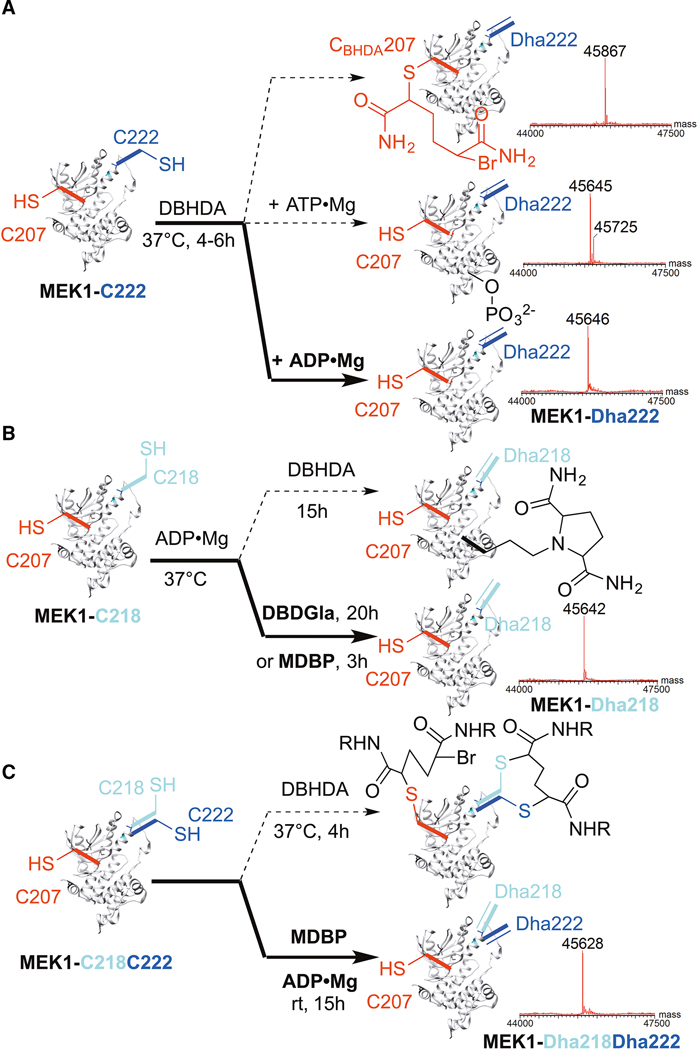
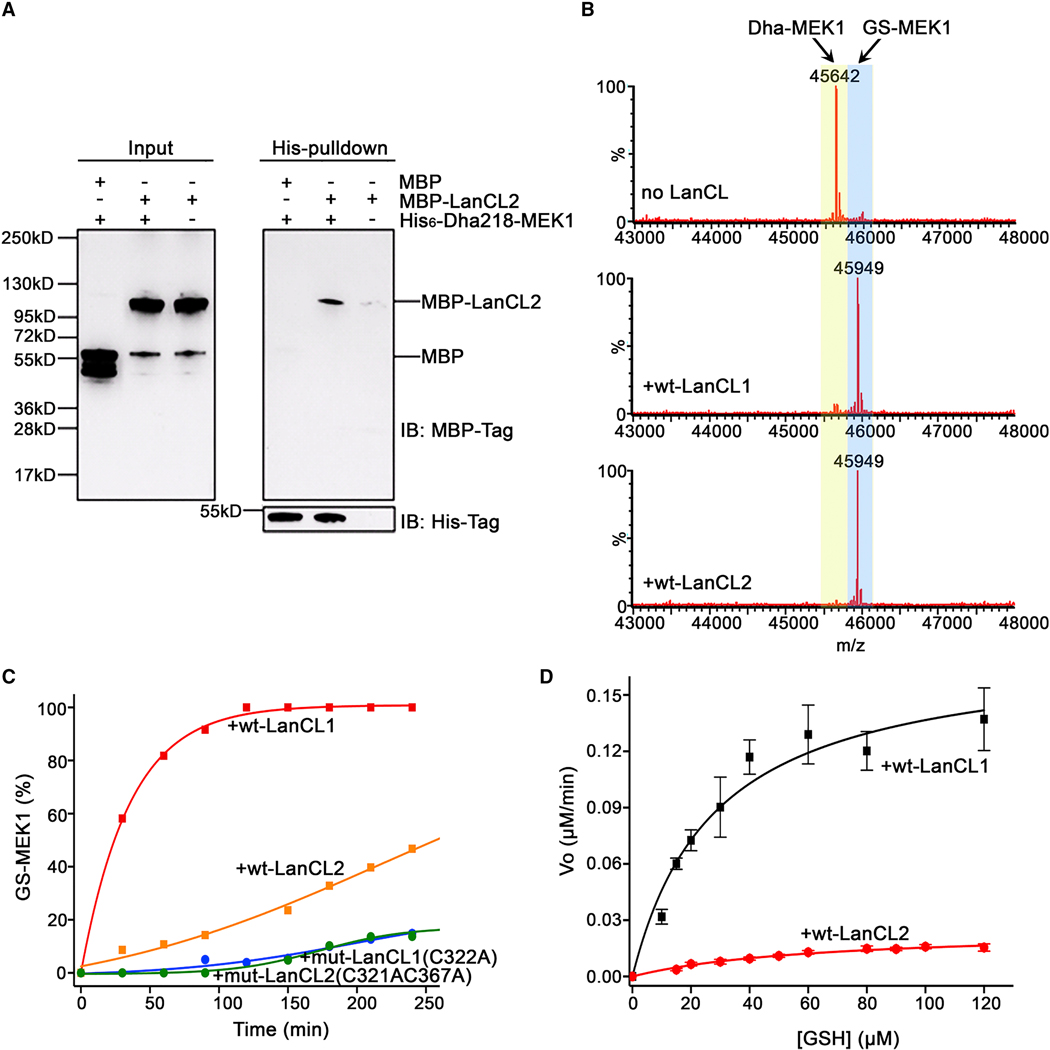
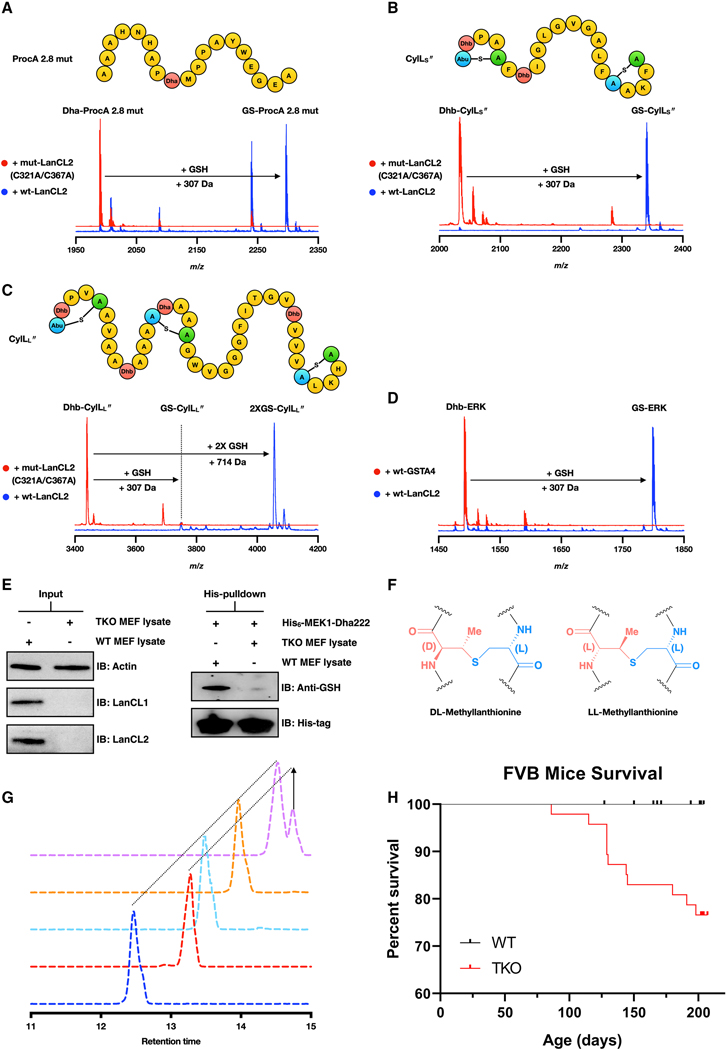
References
-
- Arbibe L, Kim DW, Batsche E, Pedron T, Mateescu B, Muchardt C, Parsot C, and Sansonetti PJ (2007). An injected bacterial effector targets chromatin access for transcription factor NF-kappaB to alter transcription of host genes involved in immune responses. Nat Immunol 8, 47–56. - PubMed
-
- Bauer H, Mayer H, Marchler-Bauer A, Salzer U, and Prohaska R. (2000). Characterization of p40/GPR69A as a peripheral membrane protein related to the lantibiotic synthetase component C. Biochem Biophys Res Commun 275, 69–74. - PubMed
-
- Brennan DF, and Barford D. (2009). Eliminylation: a post-translational modification catalyzed by phosphothreonine lyases. Trends Biochem Sci 34, 108–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
