

Regulation of hippocamposeptal input within the medial septum/diagonal band of Broca
- PMID: 33933476
- PMCID: PMC8217288
- DOI: 10.1016/j.neuropharm.2021.108589
Regulation of hippocamposeptal input within the medial septum/diagonal band of Broca
Abstract
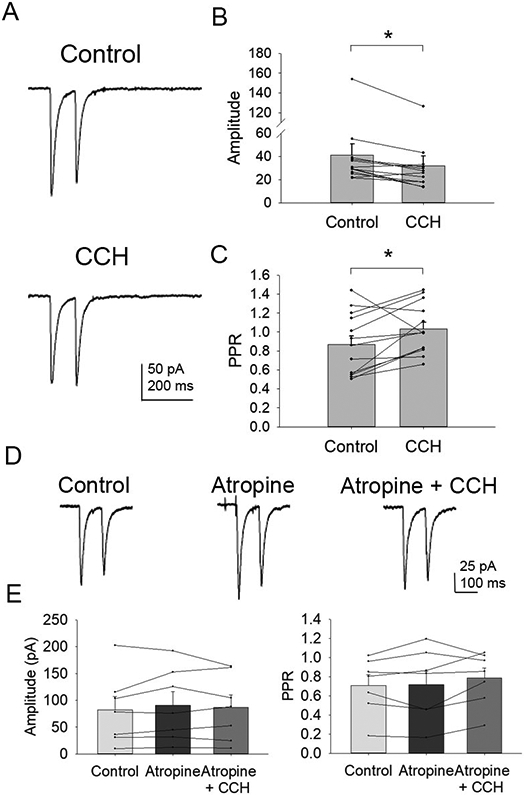
The medial septum/diagonal band of Broca (MS/DBB) receives direct GABAergic input from the hippocampus via hippocamposeptal (HS) projection neurons as part of a reciprocal loop that mediates cognition and is altered in Alzheimer's disease. Cholinergic and GABAergic interactions occur throughout the MS/DBB, but it is not known how HS GABA release is impacted by these circuits. Most HS neurons contain somatostatin (SST), so to evoke HS GABA release we expressed Cre-dependent mCherry/channelrhodopisin-2 (ChR2) in the hippocampi of SST-IRES-Cre mice and then used optogenetics to stimulate HS fibers while performing whole-cell patch clamp recordings from MS/DBB neurons in acute slices. We found that the acetylcholine receptor (AChR) agonist carbachol and the GABAB receptor (GABABR) agonist baclofen significantly decreased HS GABA release in the MS/DBB. Carbachol's effects were blocked by eliminating local GABAergic activity or inhibiting GABABRs, indicating that it was indirectly decreasing HS GABA release by increasing GABAergic tone. There was no effect of acute exposure to amyloid-β on HS GABA release. Repetitive stimulation of HS fibers increased spontaneous GABA release in the MS/DBB, revealing that HS projections can modulate local GABAergic tone. These results show that HS GABA release has far-reaching impacts on overall levels of inhibition in the MS/DBB and is under regulatory control by cholinergic and GABAergic activity. This bidirectional modulation of GABA release from local and HS projections in the MS/DBB will likely have profound impact not only on activity within the MS/DBB, but also on output to the hippocampus and hippocampal-dependent learning and memory.
Keywords: Acetylcholine; Basal forebrain; Cholinergic; GABA; Hippocamposeptal; Medial septum.
Published by Elsevier Ltd.
Conflict of interest statement
Figures







References
-
- Alonso A, Kohler C (1982) Evidence for separate projections of hippocampal pyramidal and non-pyramidal neurons to different parts of the septum in the rat brain. Neuroscience letters 31:209–214. - PubMed
-
- Alreja M, Wu M, Liu W, Atkins JB, Leranth C, Shanabrough M (2000) Muscarinic tone sustains impulse flow in the septohippocampal GABA but not cholinergic pathway: implications for learning and memory. The Journal of neuroscience : the official journal of the Society for Neuroscience 20:8103–8110. - PMC - PubMed
-
- Bartus RT, Dean RL, Beer B, Lippa AS (1982) The Cholinergic Hypothesis of Geriatric Memory Dysfunction. Science 217:408–417. - PubMed
-
- Blasco-Ibanez JM, Freund TF (1995) Synaptic input of horizontal interneurons in stratum oriens of the hippocampal CA1 subfield: structural basis of feed-back activation. The European journal of neuroscience 7:2170–2180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
