

An interpreted atlas of biosynthetic gene clusters from 1,000 fungal genomes
- PMID: 33941694
- PMCID: PMC8126772
- DOI: 10.1073/pnas.2020230118
An interpreted atlas of biosynthetic gene clusters from 1,000 fungal genomes
Abstract
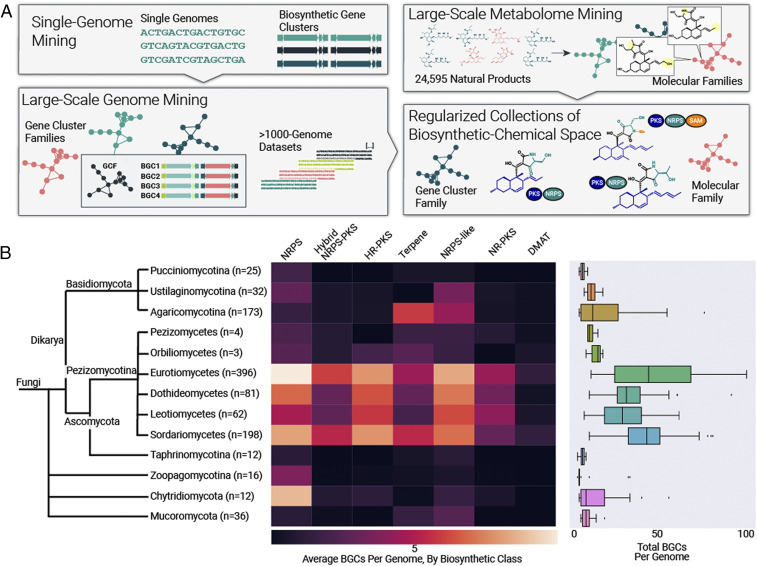
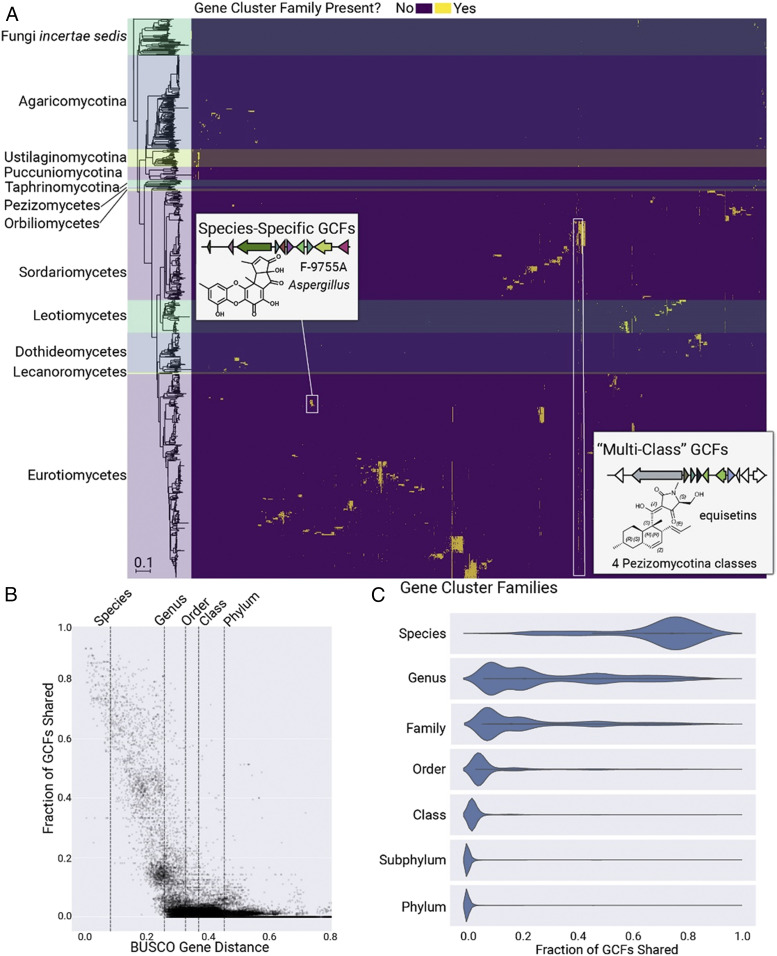
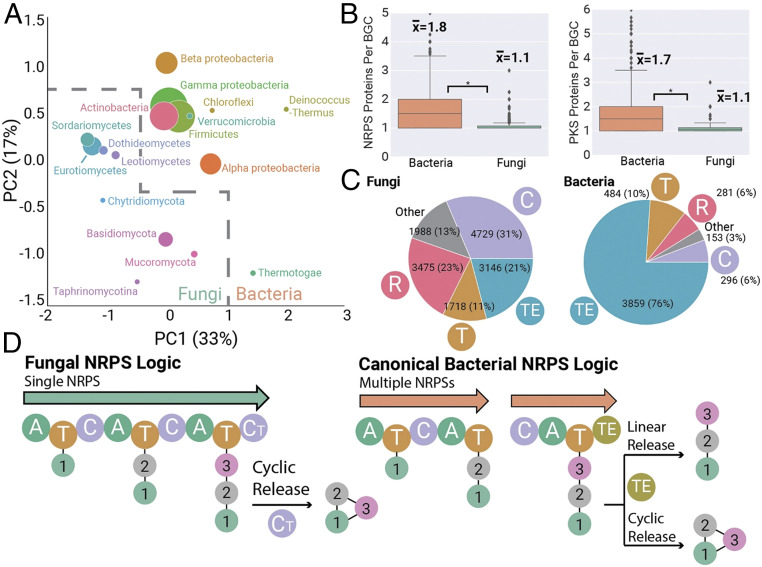
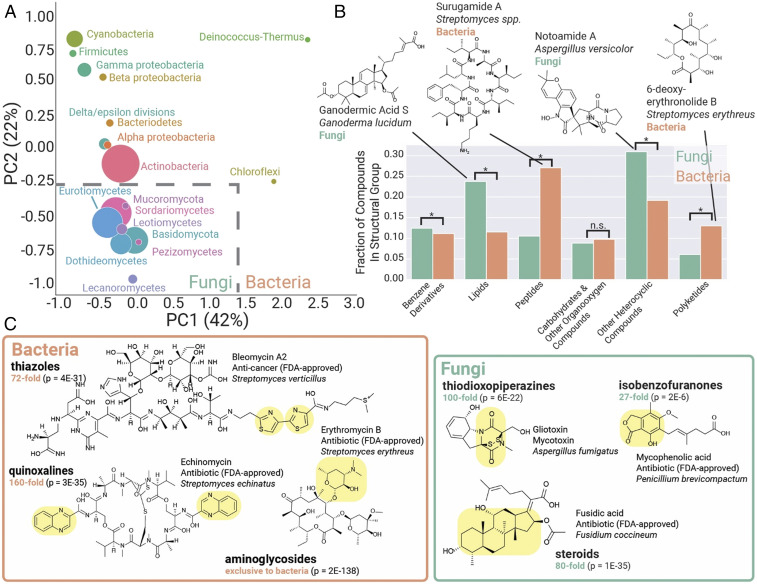
Fungi are prolific producers of natural products, compounds which have had a large societal impact as pharmaceuticals, mycotoxins, and agrochemicals. Despite the availability of over 1,000 fungal genomes and several decades of compound discovery efforts from fungi, the biosynthetic gene clusters (BGCs) encoded by these genomes and the associated chemical space have yet to be analyzed systematically. Here, we provide detailed annotation and analyses of fungal biosynthetic and chemical space to enable genome mining and discovery of fungal natural products. Using 1,037 genomes from species across the fungal kingdom (e.g., Ascomycota, Basidiomycota, and non-Dikarya taxa), 36,399 predicted BGCs were organized into a network of 12,067 gene cluster families (GCFs). Anchoring these GCFs with reference BGCs enabled automated annotation of 2,026 BGCs with predicted metabolite scaffolds. We performed parallel analyses of the chemical repertoire of fungi, organizing 15,213 fungal compounds into 2,945 molecular families (MFs). The taxonomic landscape of fungal GCFs is largely species specific, though select families such as the equisetin GCF are present across vast phylogenetic distances with parallel diversifications in the GCF and MF. We compare these fungal datasets with a set of 5,453 bacterial genomes and their BGCs and 9,382 bacterial compounds, revealing dramatic differences between bacterial and fungal biosynthetic logic and chemical space. These genomics and cheminformatics analyses reveal the large extent to which fungal and bacterial sources represent distinct compound reservoirs. With a >10-fold increase in the number of interpreted strains and annotated BGCs, this work better regularizes the biosynthetic potential of fungi for rational compound discovery.
Keywords: biosynthesis; fungi; genome mining; natural products; secondary metabolism.
Conflict of interest statement
The authors declare no competing interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
