

The voltage sensor is responsible for ΔpH dependence in Hv1 channels
- PMID: 33941706
- PMCID: PMC8126849
- DOI: 10.1073/pnas.2025556118
The voltage sensor is responsible for ΔpH dependence in Hv1 channels
Abstract
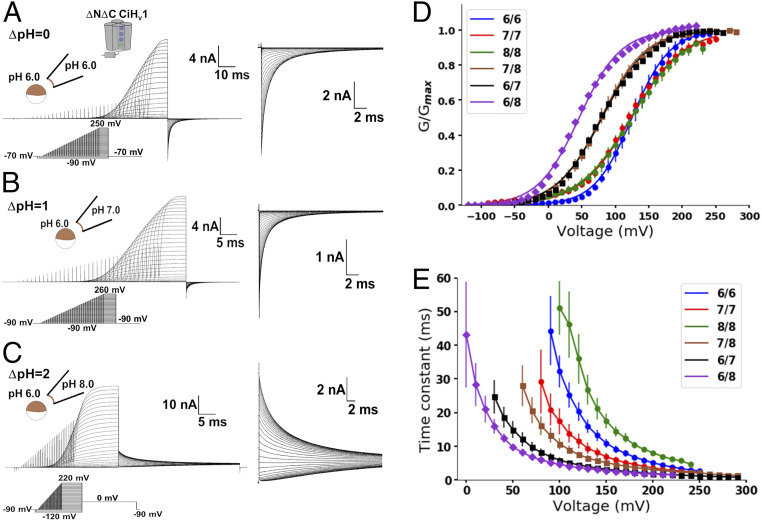
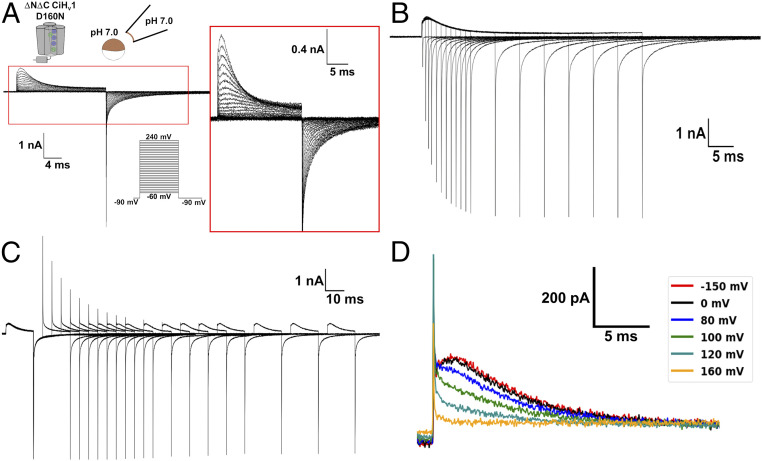
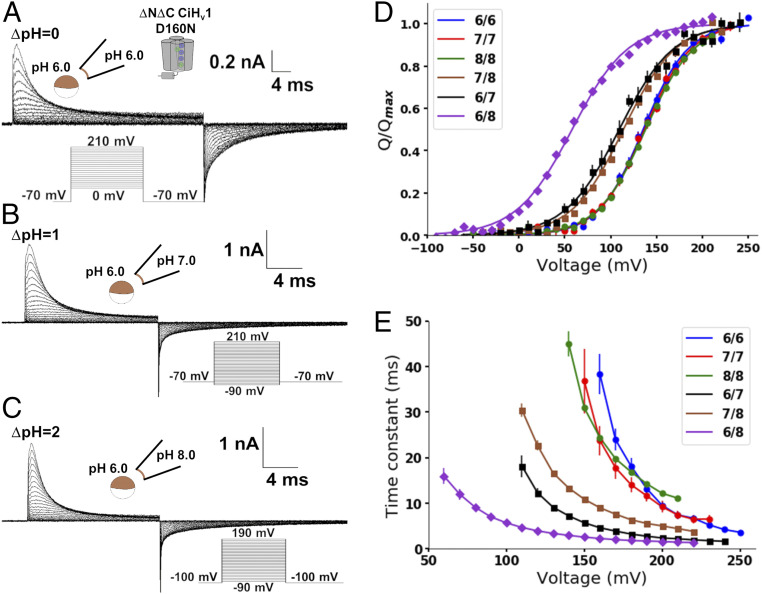
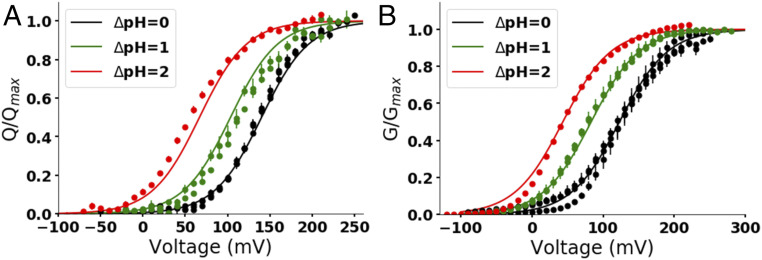
The dissipation of acute acid loads by the voltage-gated proton channel (Hv1) relies on regulating the channel's open probability by the voltage and the ΔpH across the membrane (ΔpH = pHex - pHin). Using monomeric Ciona-Hv1, we asked whether ΔpH-dependent gating is produced during the voltage sensor activation or permeation pathway opening. A leftward shift of the conductance-voltage (G-V) curve was produced at higher ΔpH values in the monomeric channel. Next, we measured the voltage sensor pH dependence in the absence of a functional permeation pathway by recording gating currents in the monomeric nonconducting D160N mutant. Increasing the ΔpH leftward shifted the gating charge-voltage (Q-V) curve, demonstrating that the ΔpH-dependent gating in Hv1 arises by modulating its voltage sensor. We fitted our data to a model that explicitly supposes the Hv1 voltage sensor free energy is a function of both the proton chemical and the electrical potential. The parameters obtained showed that around 60% of the free energy stored in the ΔpH is coupled to the Hv1 voltage sensor activation. Our results suggest that the molecular mechanism underlying the Hv1 ΔpH dependence is produced by protons, which alter the free-energy landscape around the voltage sensor domain. We propose that this alteration is produced by accessibility changes of the protons in the Hv1 voltage sensor during activation.
Keywords: Hv1; coupling; gating currents; pH dependence; voltage sensor.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Mozhayeva G. N., Naumov A. P., Effect of surface charge on the steady-state potassium conductance of nodal membrane. Nature 228, 164–165 (1970). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
