

Multidrug Resistance in Mammals and Fungi-From MDR to PDR: A Rocky Road from Atomic Structures to Transport Mechanisms
- PMID: 33946618
- PMCID: PMC8124828
- DOI: 10.3390/ijms22094806
Multidrug Resistance in Mammals and Fungi-From MDR to PDR: A Rocky Road from Atomic Structures to Transport Mechanisms
Abstract
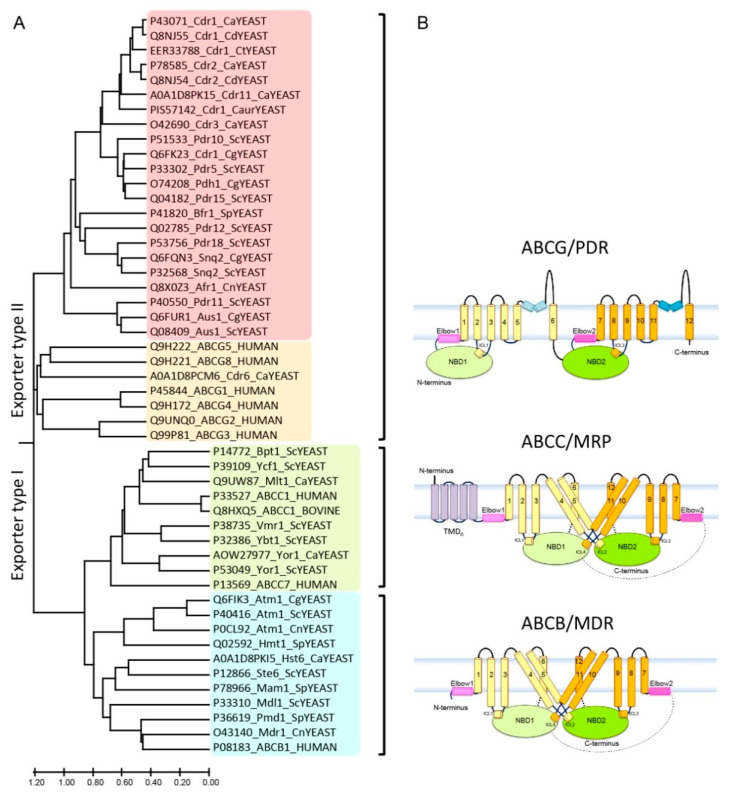
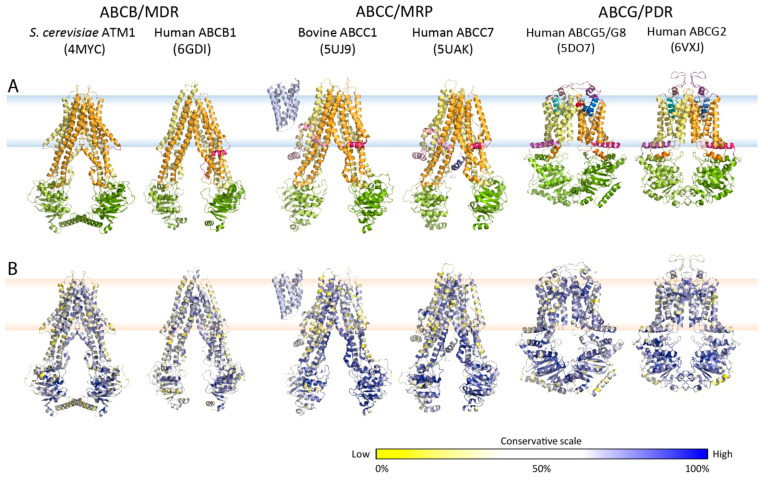
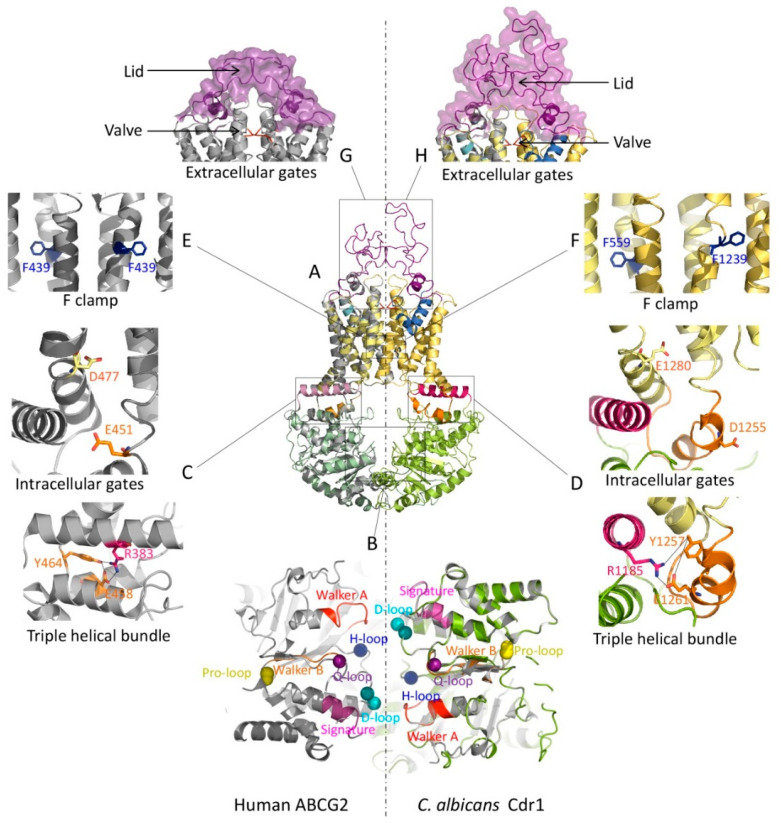
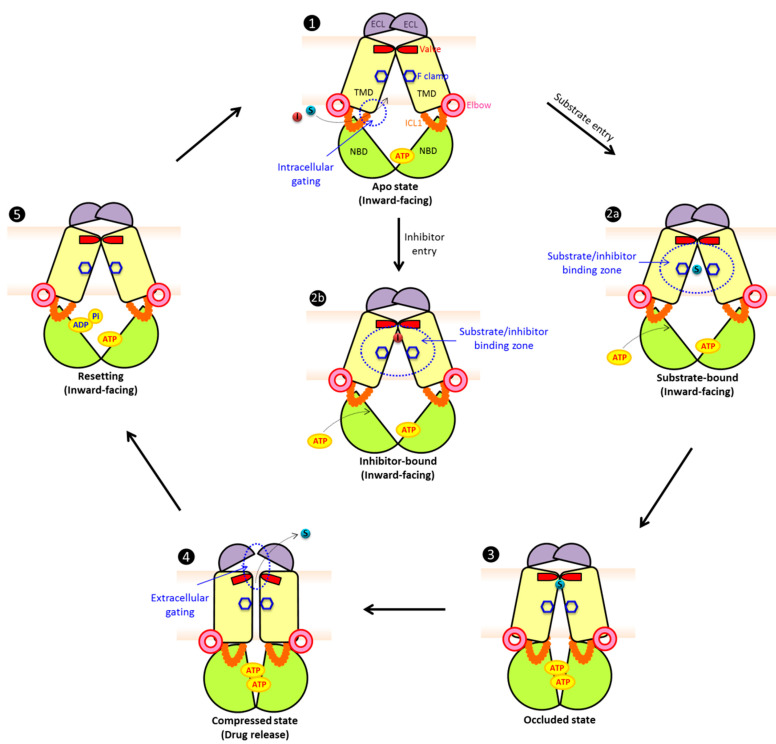
Multidrug resistance (MDR) can be a serious complication for the treatment of cancer as well as for microbial and parasitic infections. Dysregulated overexpression of several members of the ATP-binding cassette transporter families have been intimately linked to MDR phenomena. Three paradigm ABC transporter members, ABCB1 (P-gp), ABCC1 (MRP1) and ABCG2 (BCRP) appear to act as brothers in arms in promoting or causing MDR in a variety of therapeutic cancer settings. However, their molecular mechanisms of action, the basis for their broad and overlapping substrate selectivity, remains ill-posed. The rapidly increasing numbers of high-resolution atomic structures from X-ray crystallography or cryo-EM of mammalian ABC multidrug transporters initiated a new era towards a better understanding of structure-function relationships, and for the dynamics and mechanisms driving their transport cycles. In addition, the atomic structures offered new evolutionary perspectives in cases where transport systems have been structurally conserved from bacteria to humans, including the pleiotropic drug resistance (PDR) family in fungal pathogens for which high resolution structures are as yet unavailable. In this review, we will focus the discussion on comparative mechanisms of mammalian ABCG and fungal PDR transporters, owing to their close evolutionary relationships. In fact, the atomic structures of ABCG2 offer excellent models for a better understanding of fungal PDR transporters. Based on comparative structural models of ABCG transporters and fungal PDRs, we propose closely related or even conserved catalytic cycles, thus offering new therapeutic perspectives for preventing MDR in infectious disease settings.
Keywords: ABC transporters; anticancer; antifungal resistance; mechanism; multidrug transporter; yeast.
Conflict of interest statement
The authors have no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
