

Recent Advances in Effector-Triggered Immunity in Plants: New Pieces in the Puzzle Create a Different Paradigm
- PMID: 33946790
- PMCID: PMC8124997
- DOI: 10.3390/ijms22094709
Recent Advances in Effector-Triggered Immunity in Plants: New Pieces in the Puzzle Create a Different Paradigm
Abstract
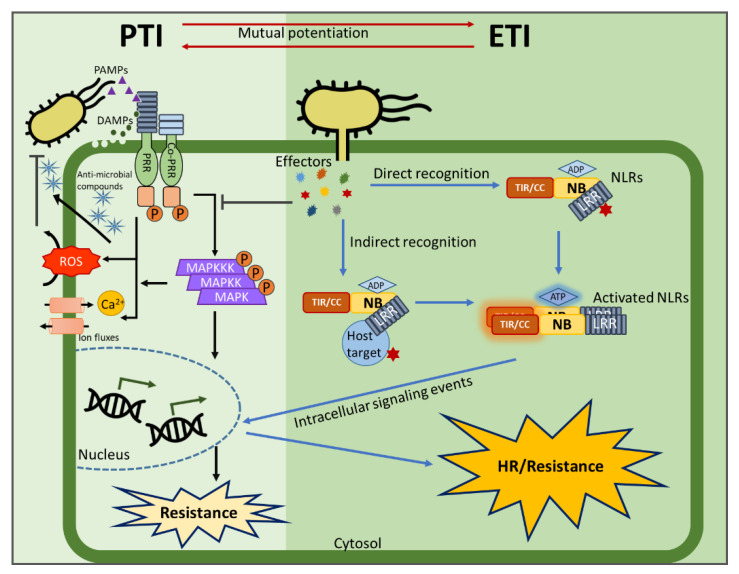
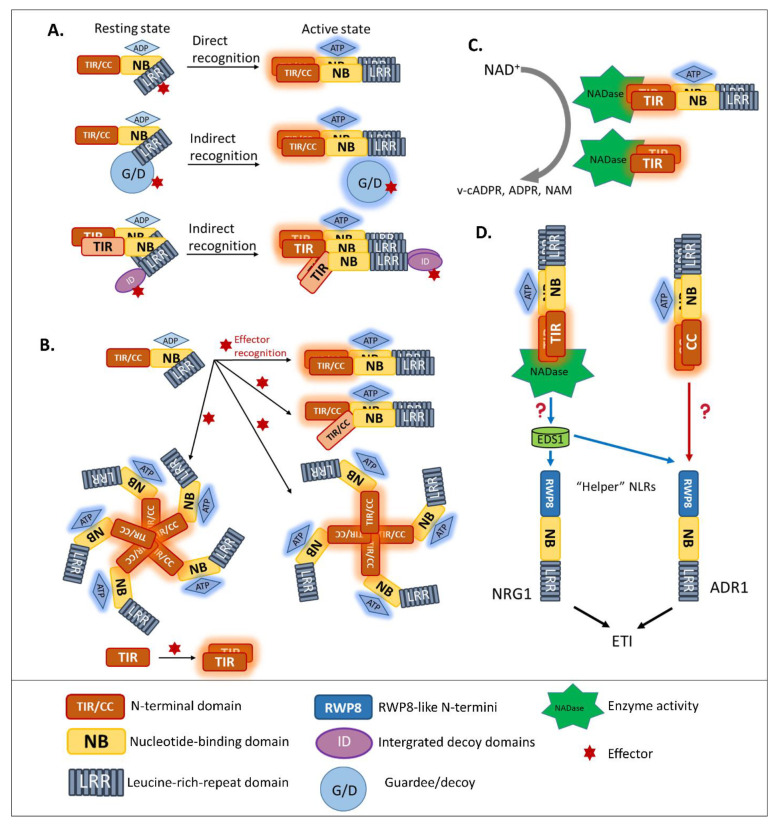
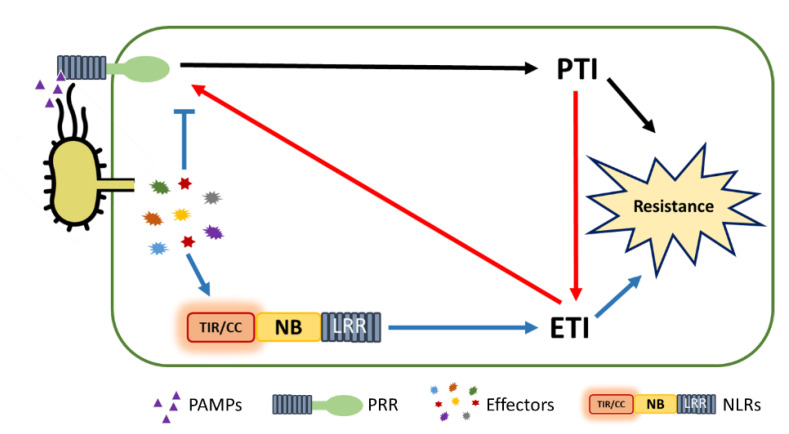
Plants rely on multiple immune systems to protect themselves from pathogens. When pattern-triggered immunity (PTI)-the first layer of the immune response-is no longer effective as a result of pathogenic effectors, effector-triggered immunity (ETI) often provides resistance. In ETI, host plants directly or indirectly perceive pathogen effectors via resistance proteins and launch a more robust and rapid defense response. Resistance proteins are typically found in the form of nucleotide-binding and leucine-rich-repeat-containing receptors (NLRs). Upon effector recognition, an NLR undergoes structural change and associates with other NLRs. The dimerization or oligomerization of NLRs signals to downstream components, activates "helper" NLRs, and culminates in the ETI response. Originally, PTI was thought to contribute little to ETI. However, most recent studies revealed crosstalk and cooperation between ETI and PTI. Here, we summarize recent advancements in our understanding of the ETI response and its components, as well as how these components cooperate in the innate immune signaling pathways. Based on up-to-date accumulated knowledge, this review provides our current perspective of potential engineering strategies for crop protection.
Keywords: ETI; NLR; PTI; effector; pathogen; plant immunity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
