

Increasing lipid yield in Yarrowia lipolytica through phosphoketolase and phosphotransacetylase expression in a phosphofructokinase deletion strain
- PMID: 33947437
- PMCID: PMC8094482
- DOI: 10.1186/s13068-021-01962-6
Increasing lipid yield in Yarrowia lipolytica through phosphoketolase and phosphotransacetylase expression in a phosphofructokinase deletion strain
Abstract
Background: Lipids are important precursors in the biofuel and oleochemical industries. Yarrowia lipolytica is among the most extensively studied oleaginous microorganisms and has been a focus of metabolic engineering to improve lipid production. Yield improvement, through rewiring of the central carbon metabolism of Y. lipolytica from glucose to the lipid precursor acetyl-CoA, is a key strategy for achieving commercial success in this organism.
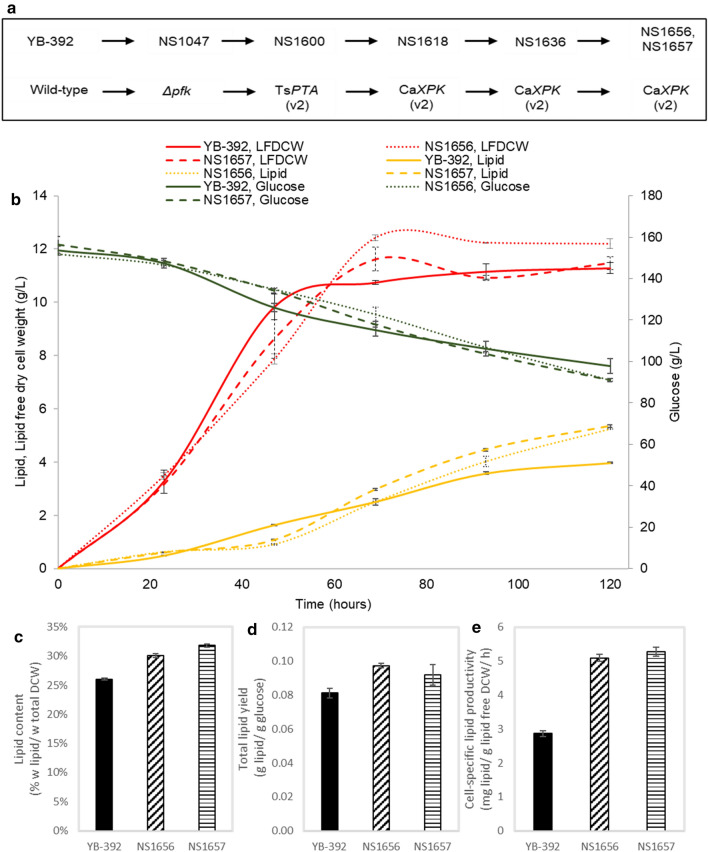
Results: Building on YB-392, a Y. lipolytica isolate known for stable non-hyphal growth and low citrate production with demonstrated potential for high lipid accumulation, we assembled a heterologous pathway that redirects carbon flux from glucose through the pentose phosphate pathway (PPP) to acetyl-CoA. We used phosphofructokinase (Pfk) deletion to block glycolysis and expressed two non-native enzymes, phosphoketolase (Xpk) and phosphotransacetylase (Pta), to convert PPP-produced xylulose-5-P to acetyl-CoA. Introduction of the pathway in a pfk deletion strain that is unable to grow and accumulate lipid from glucose in defined media ensured maximal redirection of carbon flux through Xpk/Pta. Expression of Xpk and Pta restored growth and lipid production from glucose. In 1-L bioreactors, the engineered strains recorded improved lipid yield and cell-specific productivity by up to 19 and 78%, respectively.
Conclusions: Yields and cell-specific productivities are important bioprocess parameters for large-scale lipid fermentations. Improving these parameters by engineering the Xpk/Pta pathway is an important step towards developing Y. lipolytica as an industrially preferred microbial biocatalyst for lipid production.
Keywords: Cell-specific lipid productivity; Central carbon metabolism; Lipid yield; Phosphoketolase; Phosphotransacetylase; Yarrowia lipolytica.
Conflict of interest statement
AK, ALC, KM, SC, GC and VT are current employees of Ginkgo Bioworks which has a commercial interest in the strains described in this study and is applying for patents on the work described in this study. The authors declare no other non-financial competing interests.
Figures





Similar articles
-
Engineering Yarrowia lipolytica for the production of β-carotene by carbon and redox rebalancing.J Biol Eng. 2025 Jan 15;19(1):6. doi: 10.1186/s13036-025-00476-1. J Biol Eng. 2025. PMID: 39815368 Free PMC article.
-
The oxidative pentose phosphate pathway is the primary source of NADPH for lipid overproduction from glucose in Yarrowia lipolytica.Metab Eng. 2015 Jul;30:27-39. doi: 10.1016/j.ymben.2015.02.007. Epub 2015 Mar 6. Metab Eng. 2015. PMID: 25747307
-
Engineering of a high lipid producing Yarrowia lipolytica strain.Biotechnol Biofuels. 2016 Mar 31;9:77. doi: 10.1186/s13068-016-0492-3. eCollection 2016. Biotechnol Biofuels. 2016. PMID: 27034715 Free PMC article.
-
Advanced Strategies for the Synthesis of Terpenoids in Yarrowia lipolytica.J Agric Food Chem. 2021 Mar 3;69(8):2367-2381. doi: 10.1021/acs.jafc.1c00350. Epub 2021 Feb 17. J Agric Food Chem. 2021. PMID: 33595318 Review.
-
Metabolic engineering in the host Yarrowia lipolytica.Metab Eng. 2018 Nov;50:192-208. doi: 10.1016/j.ymben.2018.07.016. Epub 2018 Jul 26. Metab Eng. 2018. PMID: 30056205 Review.
Cited by
-
Engineering a Phosphoketolase Pathway to Supplement Cytosolic Acetyl-CoA in Aspergillus niger Enables a Significant Increase in Citric Acid Production.J Fungi (Basel). 2023 Apr 23;9(5):504. doi: 10.3390/jof9050504. J Fungi (Basel). 2023. PMID: 37233215 Free PMC article.
-
The role of ATP citrate lyase, phosphoketolase, and malic enzyme in oleaginous Rhodotorula toruloides.Appl Microbiol Biotechnol. 2025 Mar 29;109(1):77. doi: 10.1007/s00253-025-13454-w. Appl Microbiol Biotechnol. 2025. PMID: 40156749 Free PMC article.
-
Advances in the optimization of central carbon metabolism in metabolic engineering.Microb Cell Fact. 2023 Apr 21;22(1):76. doi: 10.1186/s12934-023-02090-6. Microb Cell Fact. 2023. PMID: 37085866 Free PMC article. Review.
-
Metabolic Engineering of Pichia pastoris for the Production of Triacetic Acid Lactone.J Fungi (Basel). 2023 Apr 20;9(4):494. doi: 10.3390/jof9040494. J Fungi (Basel). 2023. PMID: 37108948 Free PMC article.
-
Exploring Yeast Diversity to Produce Lipid-Based Biofuels from Agro-Forestry and Industrial Organic Residues.J Fungi (Basel). 2022 Jun 29;8(7):687. doi: 10.3390/jof8070687. J Fungi (Basel). 2022. PMID: 35887443 Free PMC article. Review.
References
-
- Donot F, Fontana A, Baccou JC, Strub C, Schorr-Galindo S. Single cell oils (SCOs) from oleaginous yeasts and moulds: Production and genetics. Biomass Bioenerg. 2014;68:135–150.
-
- Koh LP, Wilcove DS. Is oil palm agriculture really destroying tropical biodiversity? Conserv Lett. 2008;1(2):60–64.
-
- Beopoulos A, Cescut J, Haddouche R, Uribelarrea J-L, Molina-Jouve C, Nicaud J-M. Yarrowia lipolytica as a model for bio-oil production. Prog Lipid Res. 2009;48(6):375–387. - PubMed
-
- Pinzi S, Pilardorado M. 4—Feedstocks for advanced biodiesel production. In: Luque R, Melero JA, editors. Advances in biodiesel production. Elsevier: Woodhead Publishing; 2012. pp. 69–90.
LinkOut - more resources
Full Text Sources
Other Literature Sources
