

A Streptococcus Quorum Sensing System Enables Suppression of Innate Immunity
- PMID: 33947757
- PMCID: PMC8262891
- DOI: 10.1128/mBio.03400-20
A Streptococcus Quorum Sensing System Enables Suppression of Innate Immunity
Abstract
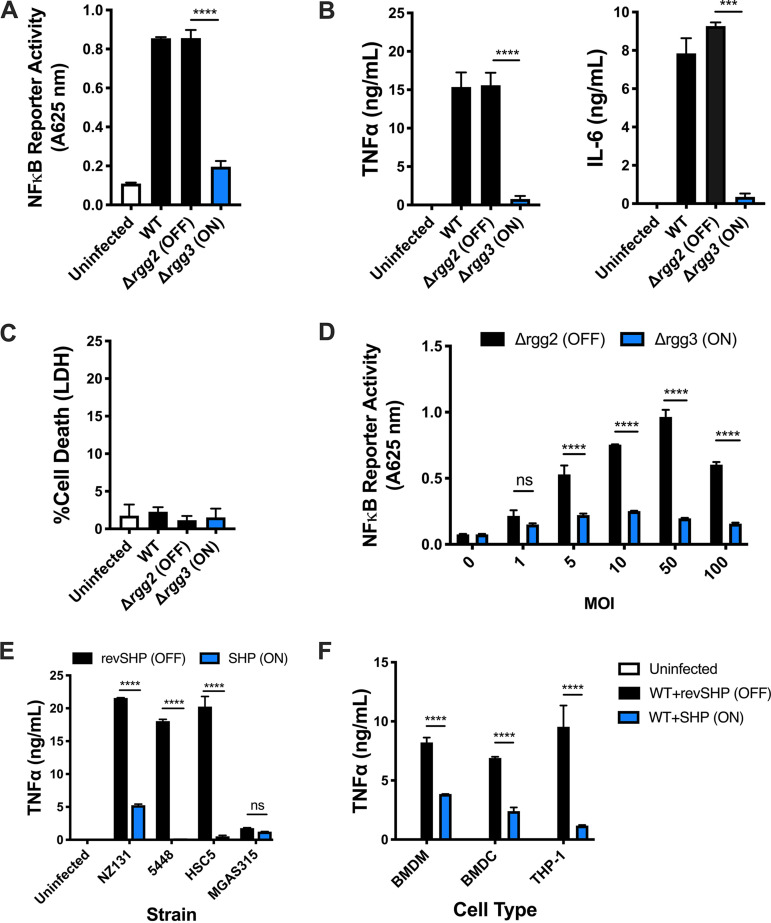
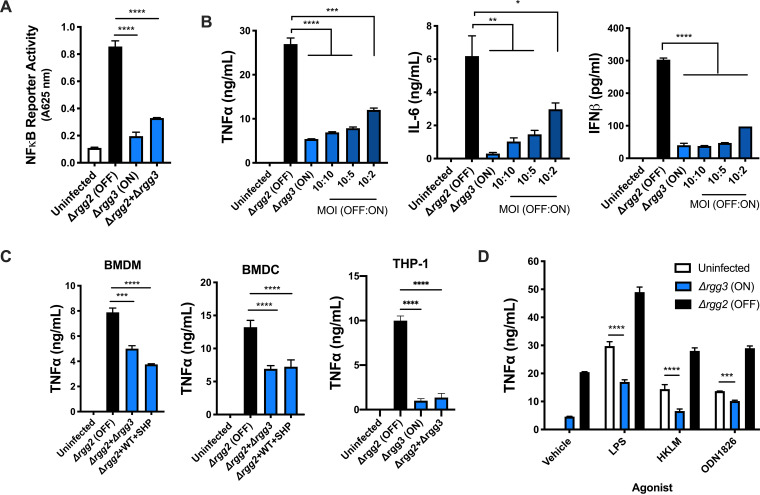
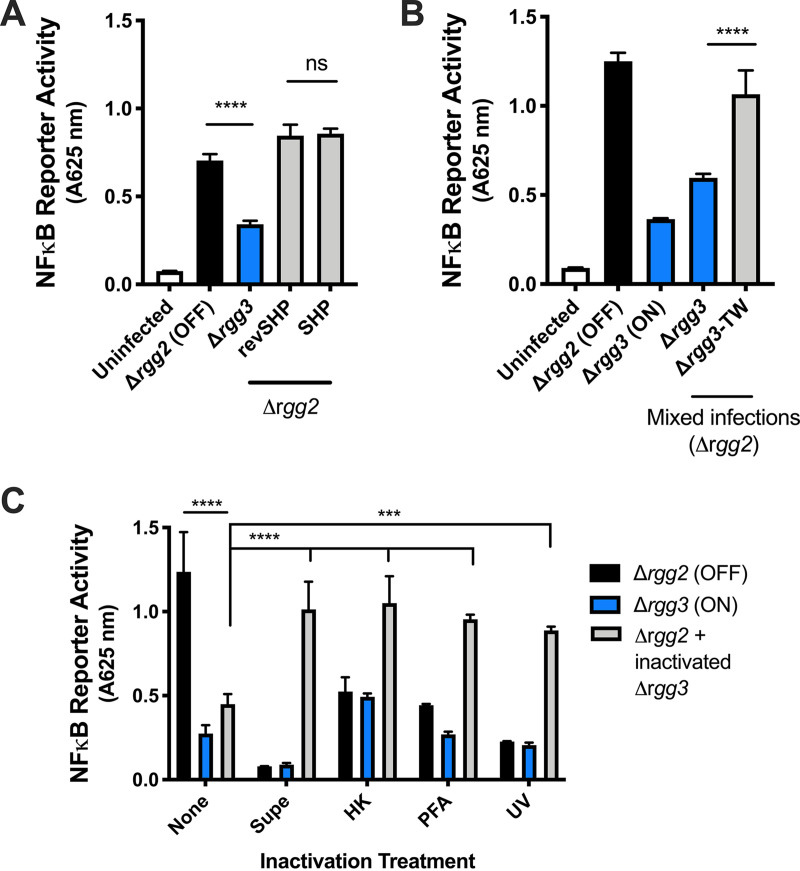
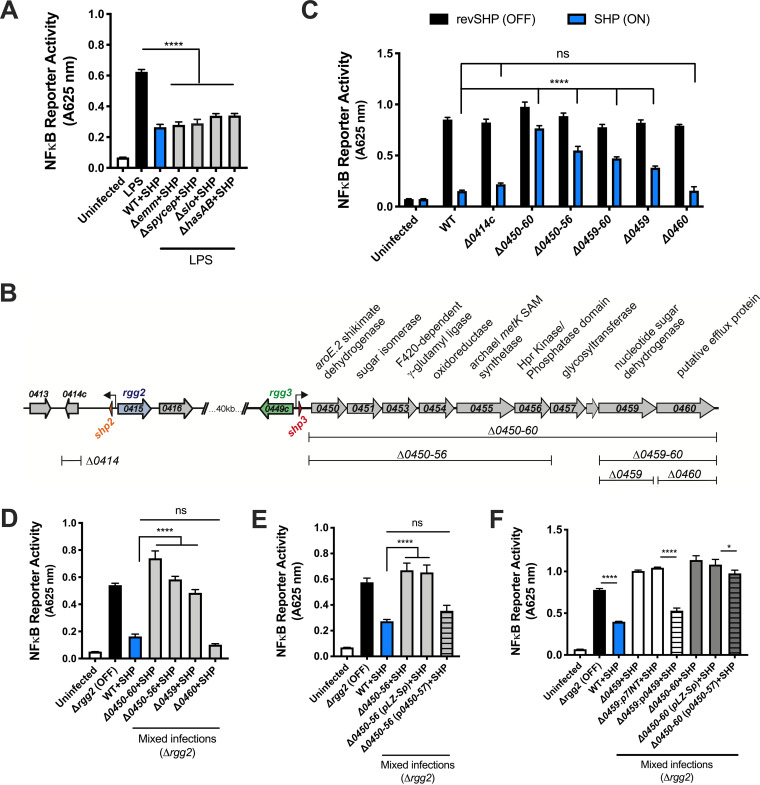
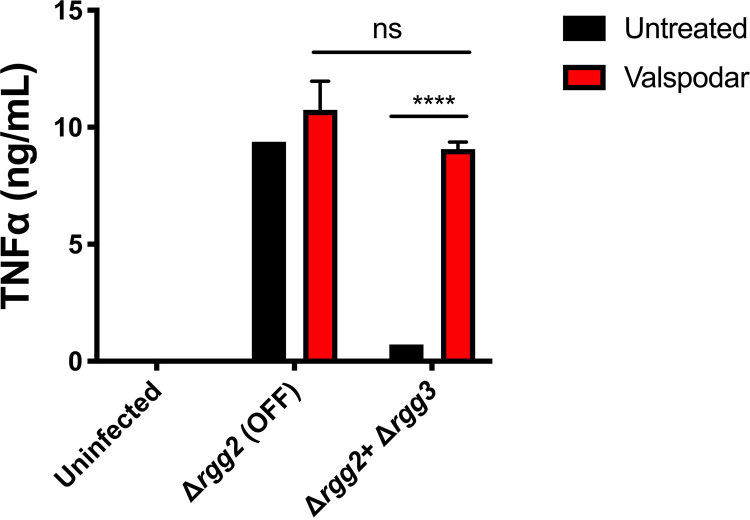
Some bacterial pathogens utilize cell-cell communication systems, such as quorum sensing (QS), to coordinate genetic programs during host colonization and infection. The human-restricted pathosymbiont Streptococcus pyogenes (group A streptococcus [GAS]) uses the Rgg2/Rgg3 QS system to modify the bacterial surface, enabling biofilm formation and lysozyme resistance. Here, we demonstrate that innate immune cell responses to GAS are substantially altered by the QS status of the bacteria. We found that macrophage activation, stimulated by multiple agonists and assessed by cytokine production and NF-κB activity, was substantially suppressed upon interaction with QS-active GAS but not QS-inactive bacteria. Neither macrophage viability nor bacterial adherence, internalization, or survival were altered by the QS activation status, yet tumor necrosis factor alpha (TNF-α), interleukin 6 (IL-6), and interferon beta (IFN-β) levels and NF-κB reporter activity were drastically lower following infection with QS-active GAS. Suppression required contact between viable bacteria and macrophages. A QS-regulated biosynthetic gene cluster (BGC) in the GAS genome, encoding several putative enzymes, was also required for macrophage modulation. Our findings suggest a model wherein upon contact with macrophages, QS-active GAS produce a BGC-derived factor capable of suppressing inflammatory responses. The suppressive capability of QS-active GAS is abolished after treatment with a specific QS inhibitor. These observations suggest that interfering with the ability of bacteria to collaborate via QS can serve as a strategy to counteract microbial efforts to manipulate host defenses.IMPORTANCEStreptococcus pyogenes is restricted to human hosts and commonly causes superficial diseases such as pharyngitis; it can also cause severe and deadly manifestations including necrotizing skin disease or severe postinfectious sequelae like rheumatic heart disease. Understanding the complex mechanisms used by this pathogen to manipulate host defenses could aid in developing new therapeutics to treat infections. Here, we examine the impact of a bacterial cell-cell communication system, which is highly conserved across S. pyogenes, on host innate immune responses. We find that S. pyogenes uses this system to suppress macrophage proinflammatory cytokine responses in vitro Interference with this communication system could serve as a strategy to disarm bacteria and maintain an effective immune response.
Keywords: NF-κB; TLR; biosynthetic gene cluster; cytokines; immunosuppression; innate immunity; macrophage; macrophages; pheromone.
Copyright © 2021 Rahbari et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
