

Preclinical evaluation for engraftment of CD34+ cells gene-edited at the sickle cell disease locus in xenograft mouse and non-human primate models
- PMID: 33948577
- PMCID: PMC8080237
- DOI: 10.1016/j.xcrm.2021.100247
Preclinical evaluation for engraftment of CD34+ cells gene-edited at the sickle cell disease locus in xenograft mouse and non-human primate models
Abstract
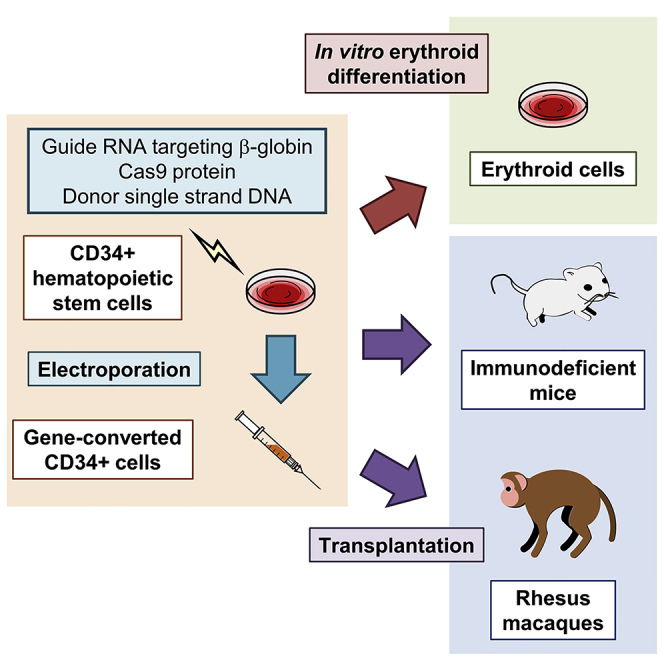
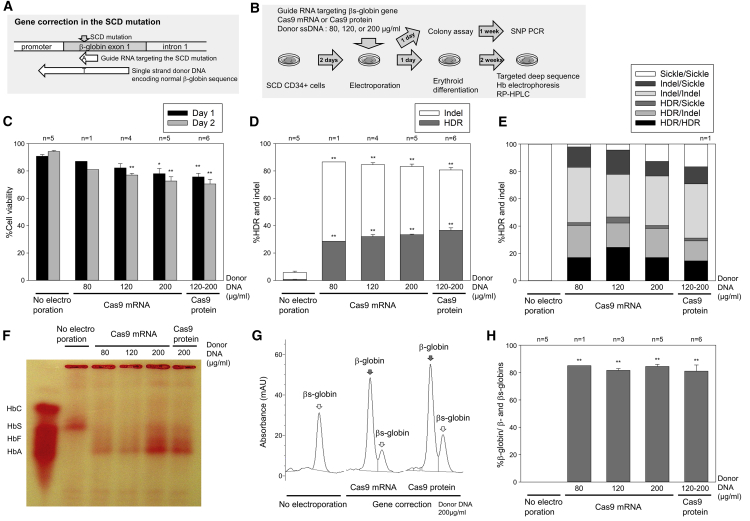
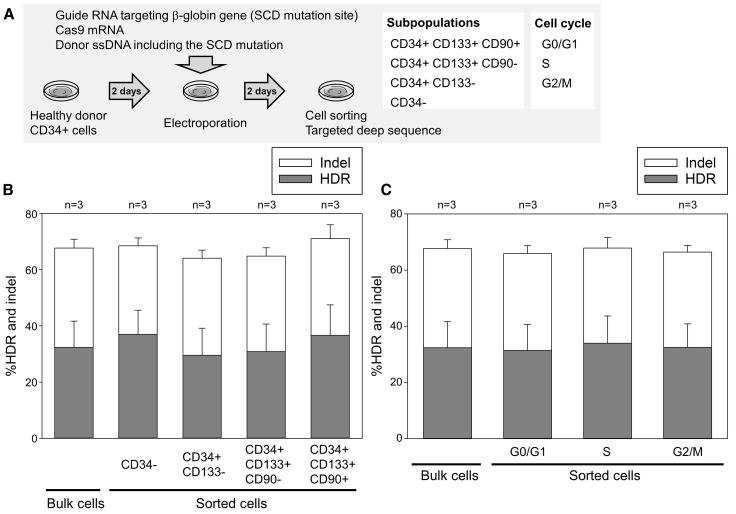
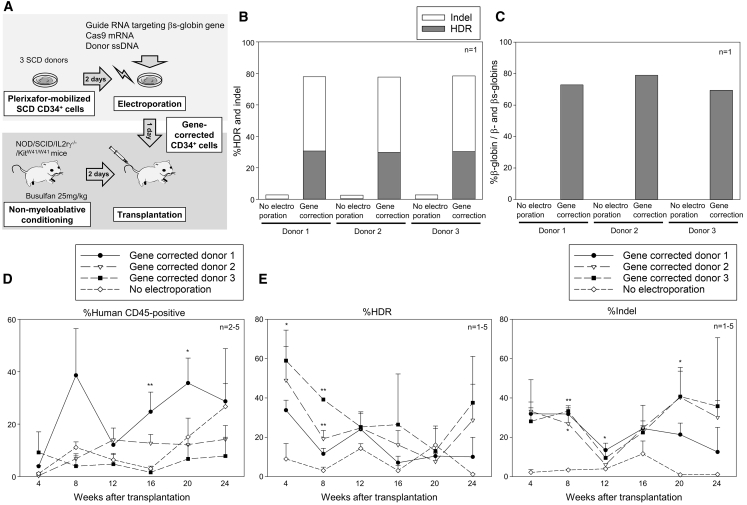
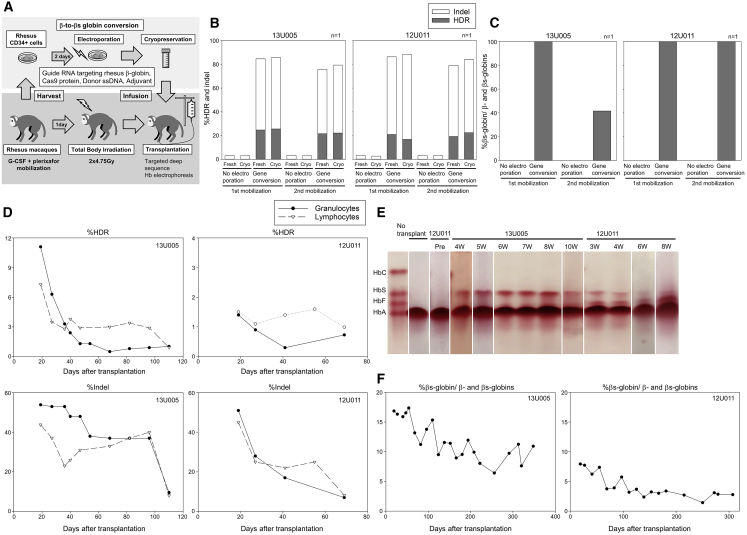
Sickle cell disease (SCD) is caused by a 20A > T mutation in the β-globin gene. Genome-editing technologies have the potential to correct the SCD mutation in hematopoietic stem cells (HSCs), producing adult hemoglobin while simultaneously eliminating sickle hemoglobin. Here, we developed high-efficiency viral vector-free non-footprint gene correction in SCD CD34+ cells with electroporation to deliver SCD mutation-targeting guide RNA, Cas9 endonuclease, and 100-mer single-strand donor DNA encoding intact β-globin sequence, achieving therapeutic-level gene correction at DNA (∼30%) and protein (∼80%) levels. Gene-edited SCD CD34+ cells contributed corrected cells 6 months post-xenograft mouse transplant without off-target δ-globin editing. We then developed a rhesus β-to-βs-globin gene conversion strategy to model HSC-targeted genome editing for SCD and demonstrate the engraftment of gene-edited CD34+ cells 10-12 months post-transplant in rhesus macaques. In summary, gene-corrected CD34+ HSCs are engraftable in xenograft mice and non-human primates. These findings are helpful in designing HSC-targeted gene correction trials.
Keywords: CRISPR/Cas9; electroporation; gene correction; genome editing; hematopoietic stem cell; large animal model; non-human primate; sickle cell disease; transplantation; β-globin gene.
Conflict of interest statement
L.L., C.A., and M.V.P. were employees at MaxCyte during the period of this work.
Figures
References
-
- Steinberg M.H. Management of sickle cell disease. N. Engl. J. Med. 1999;340:1021–1030. - PubMed
-
- Bridges K.R., Barabino G.D., Brugnara C., Cho M.R., Christoph G.W., Dover G., Ewenstein B.M., Golan D.E., Guttmann C.R., Hofrichter J. A multiparameter analysis of sickle erythrocytes in patients undergoing hydroxyurea therapy. Blood. 1996;88:4701–4710. - PubMed
-
- Niihara Y., Miller S.T., Kanter J., Lanzkron S., Smith W.R., Hsu L.L., Gordeuk V.R., Viswanathan K., Sarnaik S., Osunkwo I., Investigators of the Phase 3 Trial of l-Glutamine in Sickle Cell Disease A Phase 3 Trial of l-Glutamine in Sickle Cell Disease. N. Engl. J. Med. 2018;379:226–235. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
