

In situ cancer vaccination using lipidoid nanoparticles
- PMID: 33952519
- PMCID: PMC8099179
- DOI: 10.1126/sciadv.abf1244
In situ cancer vaccination using lipidoid nanoparticles
Abstract
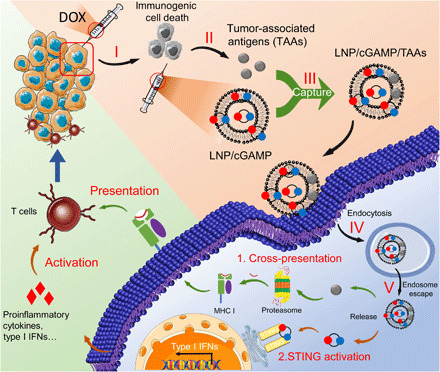
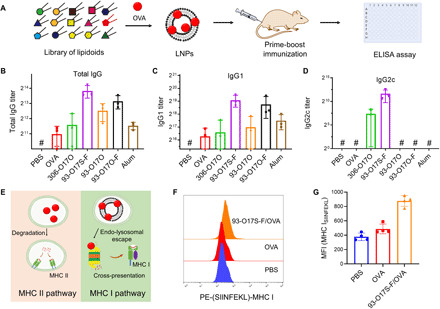
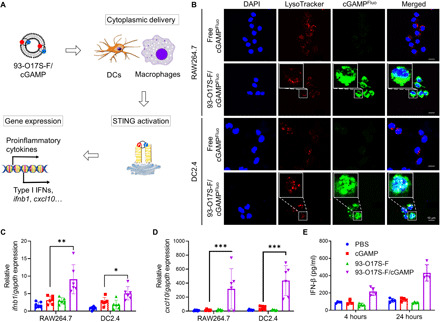
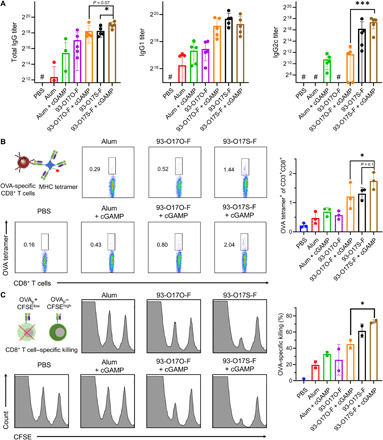
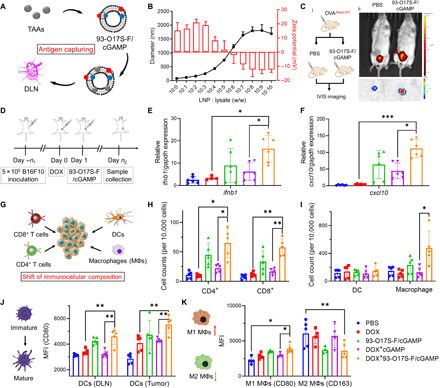
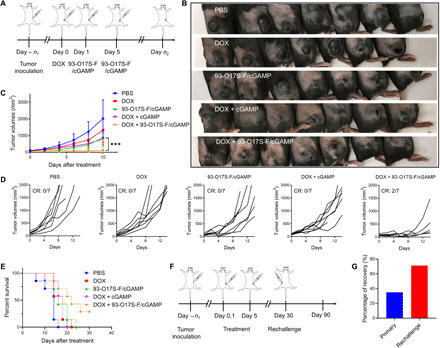
In situ vaccination is a promising strategy for cancer immunotherapy owing to its convenience and the ability to induce numerous tumor antigens. However, the advancement of in situ vaccination techniques has been hindered by low cross-presentation of tumor antigens and the immunosuppressive tumor microenvironment. To balance the safety and efficacy of in situ vaccination, we designed a lipidoid nanoparticle (LNP) to achieve simultaneously enhancing cross-presentation and STING activation. From combinatorial library screening, we identified 93-O17S-F, which promotes both the cross-presentation of tumor antigens and the intracellular delivery of cGAMP (STING agonist). Intratumor injection of 93-O17S-F/cGAMP in combination with pretreatment with doxorubicin exhibited excellent antitumor efficacy, with 35% of mice exhibiting total recovery from a primary B16F10 tumor and 71% of mice with a complete recovery from a subsequent challenge, indicating the induction of an immune memory against the tumor. This study provides a promising strategy for in situ cancer vaccination.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Kantoff P. W., Higano C. S., Shore N. D., Berger E. R., Small E. J., Penson D. F., Redfern C. H., Ferrari A. C., Dreicer R., Sims R. B., Xu Y., Frohlich M. W., Schellhammer P. F.; IMPACT Study Investigators , Sipuleucel-T immunotherapy for castration-resistant prostate cancer. N. Engl. J. Med. 363, 411–422 (2010). - PubMed
-
- Melero I., Gaudernack G., Gerritsen W., Huber C., Parmiani G., Scholl S., Thatcher N., Wagstaff J., Zielinski C., Faulkner I., Mellstedt H., Therapeutic vaccines for cancer: An overview of clinical trials. Nat. Rev. Clin. Oncol. 11, 509–524 (2014). - PubMed
-
- Martins F., Sofiya L., Sykiotis G. P., Lamine F., Maillard M., Fraga M., Shabafrouz K., Ribi C., Cairoli A., Guex-Crosier Y., Kuntzer T., Michielin O., Peters S., Coukos G., Spertini F., Thompson J. A., Obeid M., Adverse effects of immune-checkpoint inhibitors: Epidemiology, management and surveillance. Nat. Rev. Clin. Oncol. 16, 563–580 (2019). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
