

Long pentraxin PTX3 is upregulated systemically and centrally after experimental neurotrauma, but its depletion leaves unaltered sensorimotor deficits or histopathology
- PMID: 33953334
- PMCID: PMC8100171
- DOI: 10.1038/s41598-021-89032-7
Long pentraxin PTX3 is upregulated systemically and centrally after experimental neurotrauma, but its depletion leaves unaltered sensorimotor deficits or histopathology
Abstract
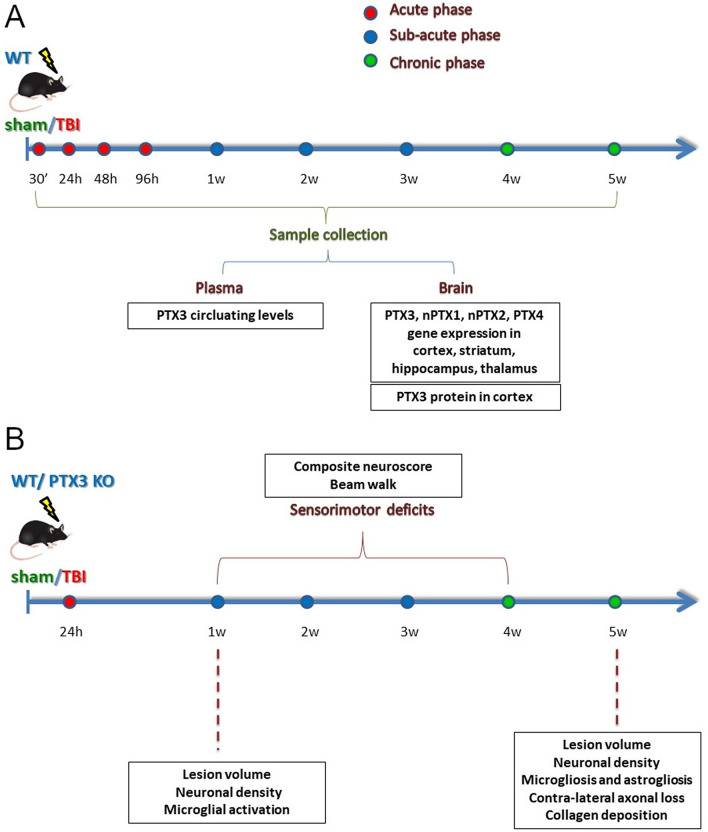
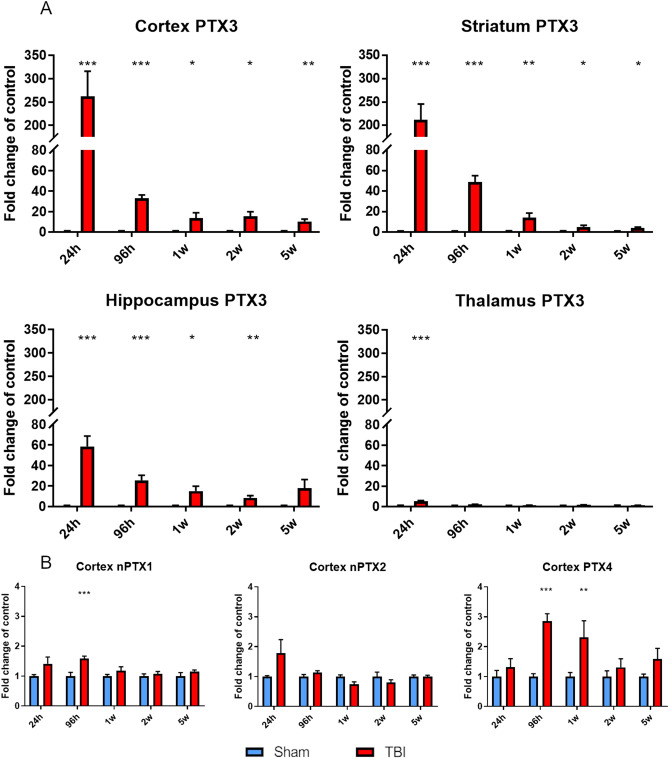
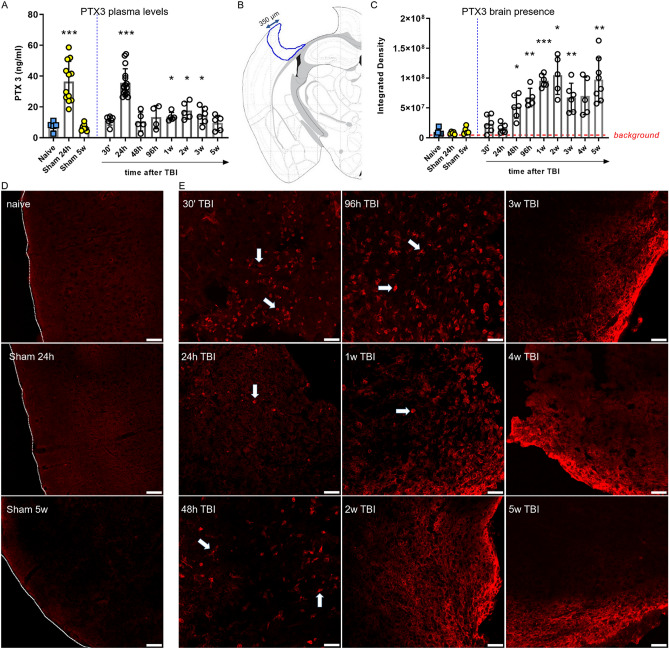
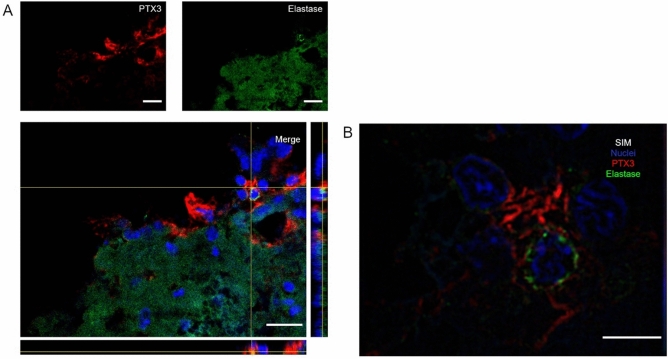
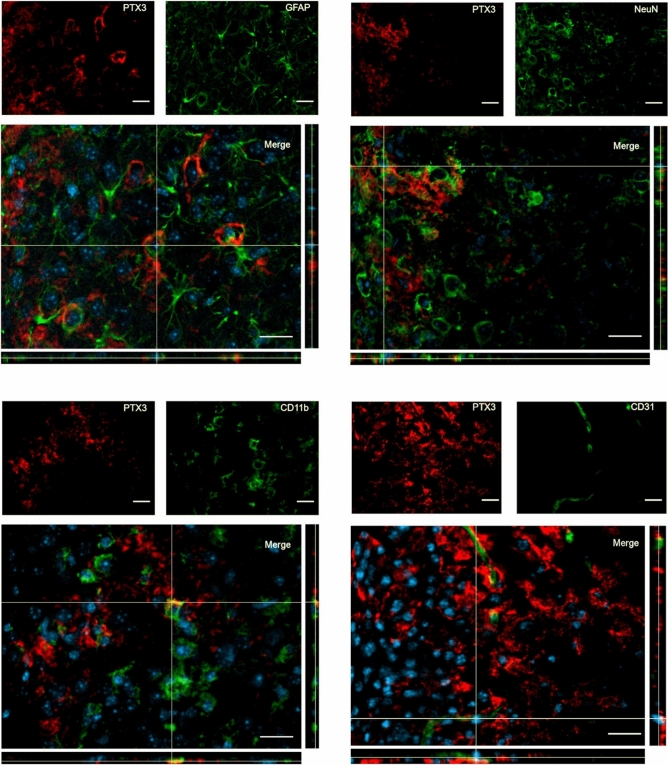
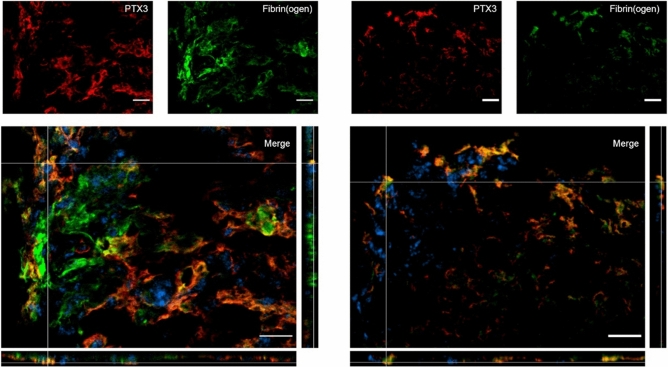
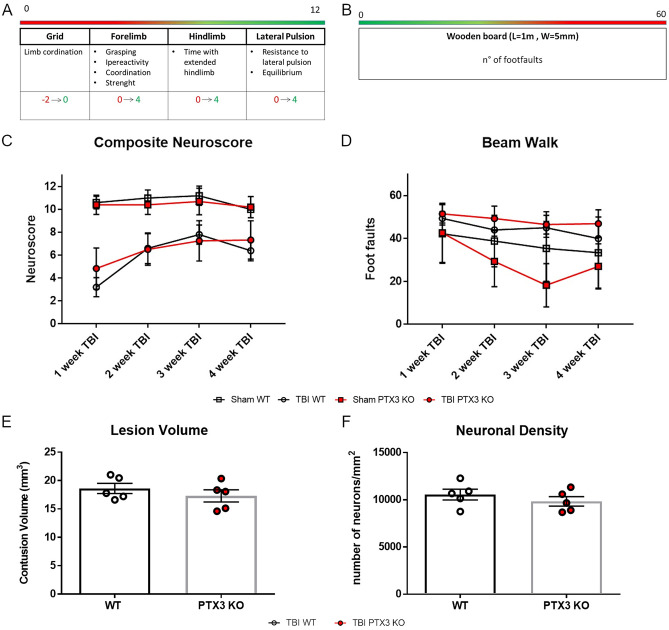
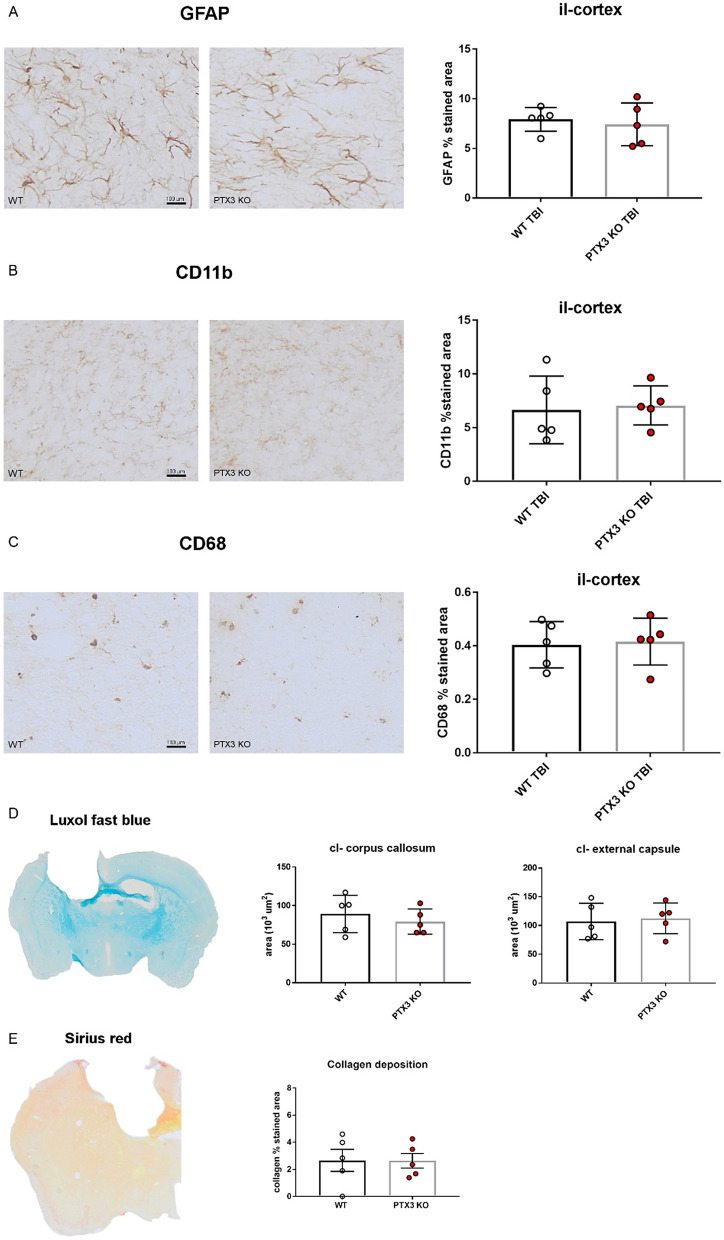
Long pentraxin PTX3, a pattern recognition molecule involved in innate immune responses, is upregulated by pro-inflammatory stimuli, contributors to secondary damage in traumatic brain injury (TBI). We analyzed PTX3 involvement in mice subjected to controlled cortical impact, a clinically relevant TBI mouse model. We measured PTX3 mRNA and protein in the brain and its circulating levels at different time point post-injury, and assessed behavioral deficits and brain damage progression in PTX3 KO mice. PTX3 circulating levels significantly increased 1-3 weeks after injury. In the brain, PTX3 mRNA was upregulated in different brain areas starting from 24 h and up to 5 weeks post-injury. PTX3 protein significantly increased in the brain cortex up to 3 weeks post-injury. Immunohistochemical analysis showed that, 48 h after TBI, PTX3 was localized in proximity of neutrophils, likely on neutrophils extracellular traps (NETs), while 1- and 2- weeks post-injury PTX3 co-localized with fibrin deposits. Genetic depletion of PTX3 did not affect sensorimotor deficits up to 5 weeks post-injury. At this time-point lesion volume and neuronal count, axonal damage, collagen deposition, astrogliosis, microglia activation and phagocytosis were not different in KO compared to WT mice. Members of the long pentraxin family, neuronal pentraxin 1 (nPTX1) and pentraxin 4 (PTX4) were also over-expressed in the traumatized brain, but not neuronal pentraxin 2 (nPTX2) or short pentraxins C-reactive protein (CRP) and serum amyloid P-component (SAP). The long-lasting pattern of activation of PTX3 in brain and blood supports its specific involvement in TBI. The lack of a clear-cut phenotype in PTX3 KO mice may depend on the different roles of this protein, possibly involved in inflammation early after injury and in repair processes later on, suggesting distinct functions in acute phases versus sub-acute or chronic phases. Brain long pentraxins, such as PTX4-shown here to be overexpressed in the brain after TBI-may compensate for PTX3 absence.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Bottazzi B, Garlanda C, Teixeira MM. Editorial: the role of pentraxins: from inflammation, tissue repair and immunity to biomarkers. Front Immunol [Internet]. 2019 [cited 2020 Jul 8];10. Available from: https://www.frontiersin.org/articles/10.3389/fimmu.2019.02817/full - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
