

Calcium-permeable AMPA receptor activity and GluA1 trafficking in the basolateral amygdala regulate operant alcohol self-administration
- PMID: 33955100
- PMCID: PMC8376775
- DOI: 10.1111/adb.13049
Calcium-permeable AMPA receptor activity and GluA1 trafficking in the basolateral amygdala regulate operant alcohol self-administration
Abstract
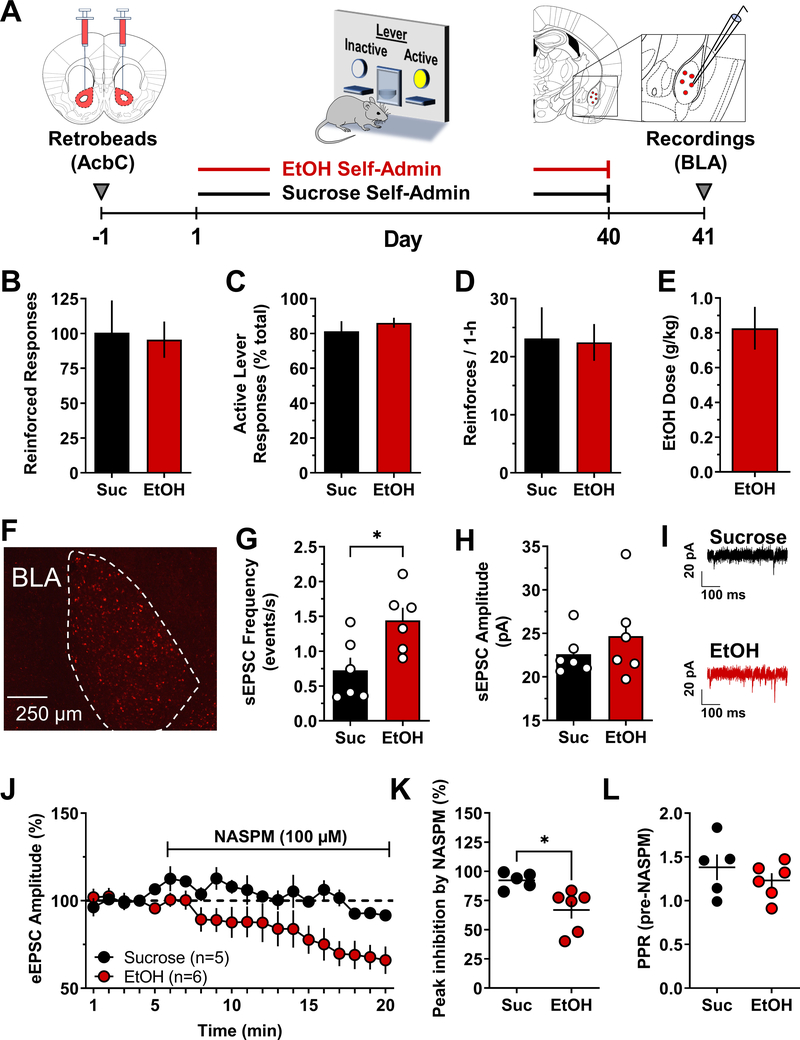
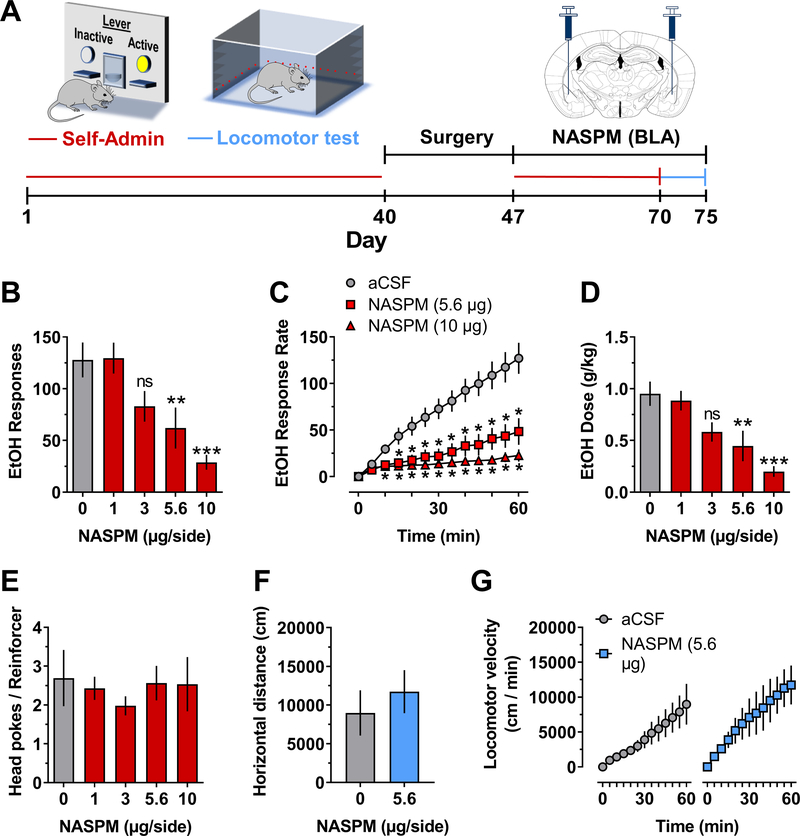
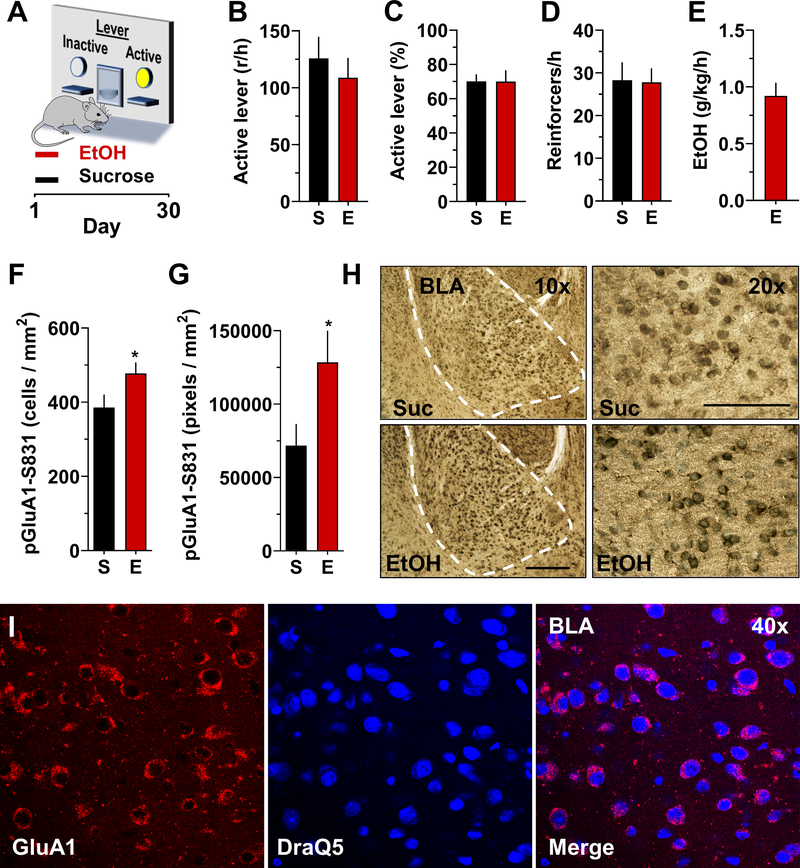
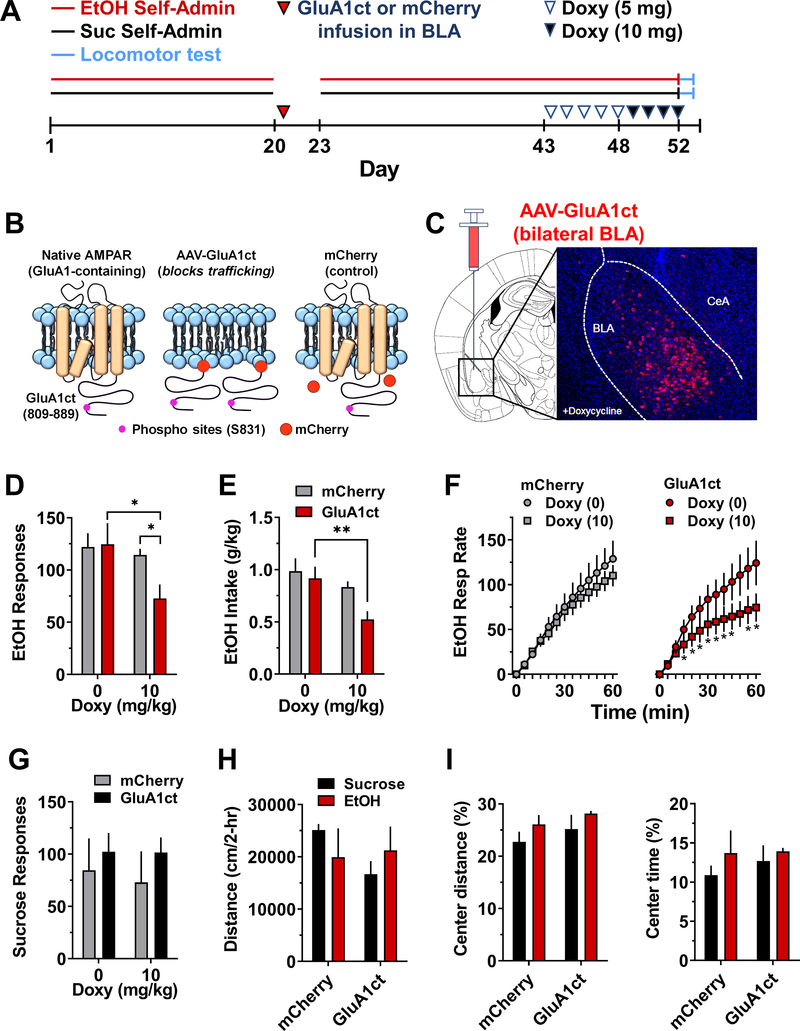
Addiction is viewed as maladaptive glutamate-mediated neuroplasticity that is regulated, in part, by calcium-permeable AMPA receptor (CP-AMPAR) activity. However, the contribution of CP-AMPARs to alcohol-seeking behavior remains to be elucidated. We evaluated CP-AMPAR activity in the basolateral amygdala (BLA) as a potential target of alcohol that also regulates alcohol self-administration in C57BL/6J mice. Operant self-administration of sweetened alcohol increased spontaneous EPSC frequency in BLA neurons that project to the nucleus accumbens as compared with behavior-matched sucrose controls indicating an alcohol-specific upregulation of synaptic activity. Bath application of the CP-AMPAR antagonist NASPM decreased evoked EPSC amplitude only in alcohol self-administering mice indicating alcohol-induced synaptic insertion of CP-AMPARs in BLA projection neurons. Moreover, NASPM infusion in the BLA dose-dependently decreased the rate of operant alcohol self-administration providing direct evidence for CP-AMPAR regulation of alcohol reinforcement. As most CP-AMPARs are GluA1-containing, we asked if alcohol alters the activation state of GluA1-containing AMPARs. Immunocytochemistry results showed elevated GluA1-S831 phosphorylation in the BLA of alcohol as compared with sucrose mice. To investigate mechanistic regulation of alcohol self-administration by GluA1-containing AMPARs, we evaluated the necessity of GluA1 trafficking using a TET-ON AAV encoding a dominant-negative GluA1 c-terminus (GluA1ct) that blocks activity-dependent synaptic delivery of native GluA1-containing AMPARs. GluA1ct expression in the BLA reduced alcohol self-administration with no effect on sucrose controls. These results show that CP-AMPAR activity and GluA1 trafficking in the BLA mechanistically regulate the reinforcing effects of sweetened alcohol. Pharmacotherapeutic targeting these mechanisms of maladaptive neuroplasticity may aid medical management of alcohol use disorder.
Keywords: GluA1; alcohol drinking; basolateral amygdala.
© 2021 Society for the Study of Addiction.
Conflict of interest statement
DISCLOSURE/CONFLICT OF INTEREST
The authors declare no conflicts of interest.
Figures
References
-
- Thorndike EL. The elements of psychology. New York:A. G. Seiler; 1905.
-
- Skinner BF. The behavior of organisms: An experimental analysis. New York: Appleton-Century-Crofts; 1938.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
