

Evidence for phloem loading via the abaxial bundle sheath cells in maize leaves
- PMID: 33955497
- PMCID: PMC8136869
- DOI: 10.1093/plcell/koaa055
Evidence for phloem loading via the abaxial bundle sheath cells in maize leaves
Abstract
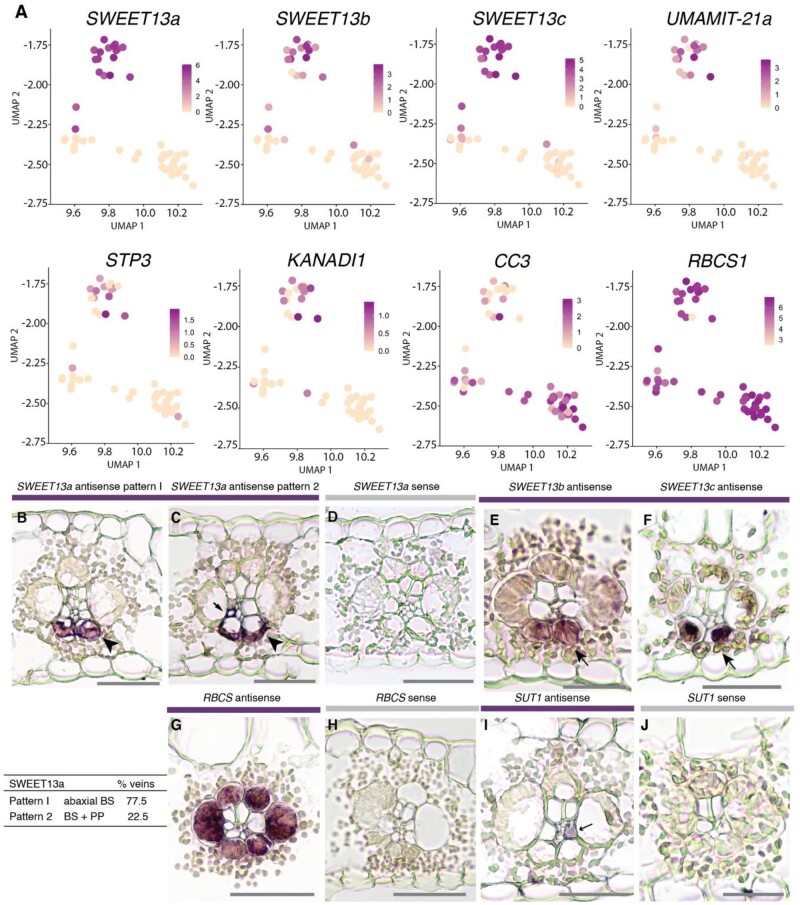
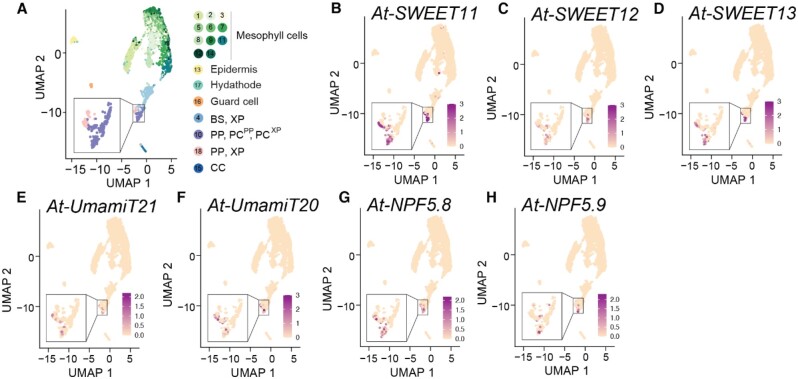
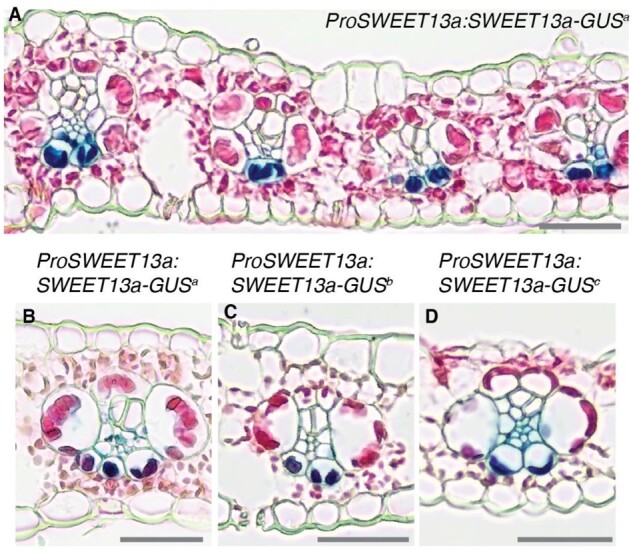
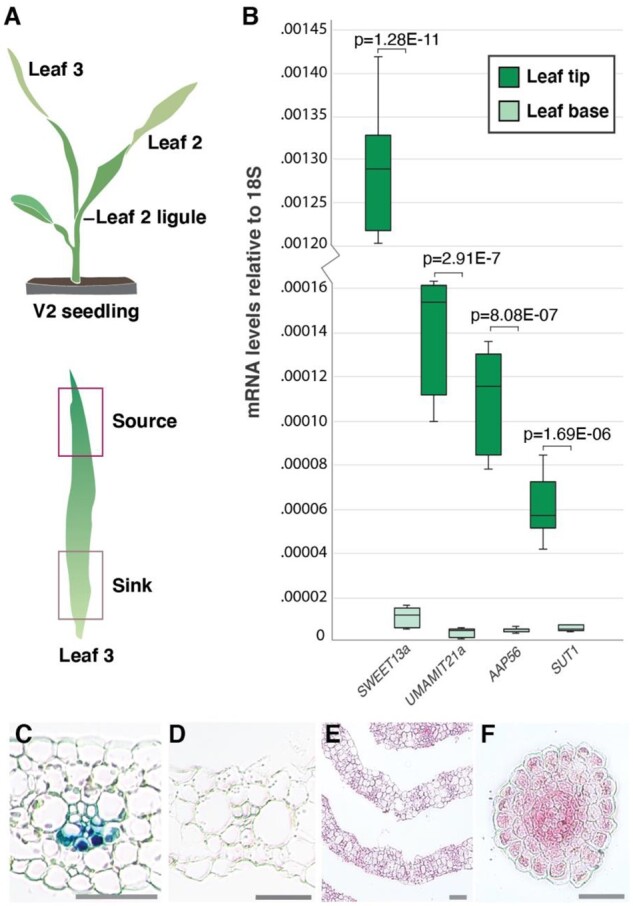
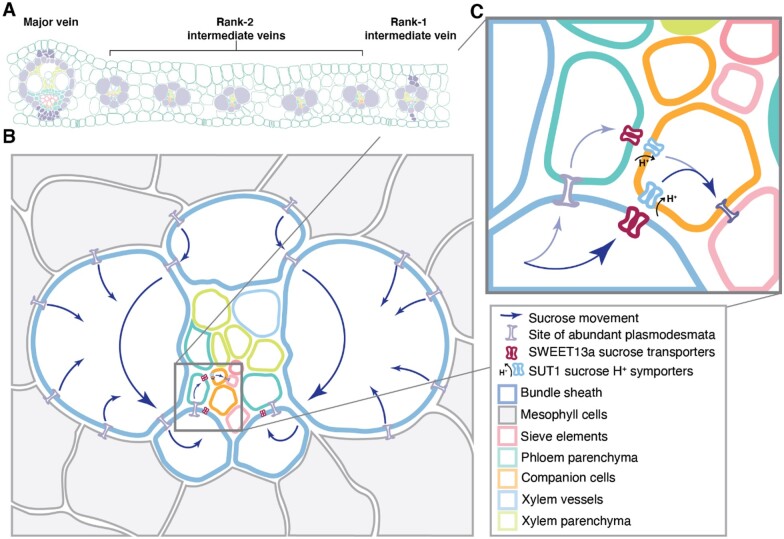
Leaves are asymmetric, with different functions for adaxial and abaxial tissue. The bundle sheath (BS) of C3 barley (Hordeum vulgare) is dorsoventrally differentiated into three types of cells: adaxial structural, lateral S-type, and abaxial L-type BS cells. Based on plasmodesmatal connections between S-type cells and mestome sheath (parenchymatous cell layer below bundle sheath), S-type cells likely transfer assimilates toward the phloem. Here, we used single-cell RNA sequencing to investigate BS differentiation in C4 maize (Zea mays L.) plants. Abaxial BS (abBS) cells of rank-2 intermediate veins specifically expressed three SWEET sucrose uniporters (SWEET13a, b, and c) and UmamiT amino acid efflux transporters. SWEET13a, b, c mRNAs were also detected in the phloem parenchyma (PP). We show that maize has acquired a mechanism for phloem loading in which abBS cells provide the main route for apoplasmic sucrose transfer toward the phloem. This putative route predominates in veins responsible for phloem loading (rank-2 intermediate), whereas rank-1 intermediate and major veins export sucrose from the PP adjacent to the sieve element companion cell complex, as in Arabidopsis thaliana. We surmise that abBS identity is subject to dorsoventral patterning and has components of PP identity. These observations provide insights into the unique transport-specific properties of abBS cells and support a modification to the canonical phloem loading pathway in maize.
© The Author(s) 2021. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures


Similar articles
-
Apoplasmic loading in the rice phloem supported by the presence of sucrose synthase and plasma membrane-localized proton pyrophosphatase.Ann Bot. 2016 Feb;117(2):257-68. doi: 10.1093/aob/mcv174. Epub 2015 Nov 26. Ann Bot. 2016. PMID: 26614751 Free PMC article.
-
Sucrose Transporter ZmSut1 Expression and Localization Uncover New Insights into Sucrose Phloem Loading.Plant Physiol. 2016 Nov;172(3):1876-1898. doi: 10.1104/pp.16.00884. Epub 2016 Sep 12. Plant Physiol. 2016. PMID: 27621426 Free PMC article.
-
Sucrose transporter1 functions in phloem loading in maize leaves.J Exp Bot. 2009;60(3):881-92. doi: 10.1093/jxb/ern335. Epub 2009 Jan 30. J Exp Bot. 2009. PMID: 19181865 Free PMC article.
-
Review: More than sweet: New insights into the biology of phloem parenchyma transfer cells in Arabidopsis.Plant Sci. 2021 Sep;310:110990. doi: 10.1016/j.plantsci.2021.110990. Epub 2021 Jul 8. Plant Sci. 2021. PMID: 34315604 Review.
-
Understanding and manipulating sucrose phloem loading, unloading, metabolism, and signalling to enhance crop yield and food security.J Exp Bot. 2014 Apr;65(7):1713-35. doi: 10.1093/jxb/ert416. Epub 2013 Dec 17. J Exp Bot. 2014. PMID: 24347463 Review.
Cited by
-
An Efficient and Universal Protoplast Isolation Protocol Suitable for Transient Gene Expression Analysis and Single-Cell RNA Sequencing.Int J Mol Sci. 2022 Mar 22;23(7):3419. doi: 10.3390/ijms23073419. Int J Mol Sci. 2022. PMID: 35408780 Free PMC article.
-
Phloem loading from below.Nat Plants. 2021 Dec;7(12):1522. doi: 10.1038/s41477-021-01065-y. Nat Plants. 2021. PMID: 34907305 No abstract available.
-
Single-cell transcriptomics: a new frontier in plant biotechnology research.Plant Cell Rep. 2024 Nov 25;43(12):294. doi: 10.1007/s00299-024-03383-9. Plant Cell Rep. 2024. PMID: 39585480 Review.
-
An RNA in situ hybridization protocol optimized for monocot tissue.STAR Protoc. 2021 Mar 17;2(2):100398. doi: 10.1016/j.xpro.2021.100398. eCollection 2021 Jun 18. STAR Protoc. 2021. PMID: 33796873 Free PMC article.
-
Advances in Single-Cell Transcriptome Sequencing and Spatial Transcriptome Sequencing in Plants.Plants (Basel). 2024 Jun 18;13(12):1679. doi: 10.3390/plants13121679. Plants (Basel). 2024. PMID: 38931111 Free PMC article. Review.
References
-
- Bezrutczyk M, Hartwig T, Horschman M, Char SN, Yang J, Yang B, Frommer WB, Sosso D (2018) Impaired phloem loading in zmsweet13a,b,c sucrose transporter triple knock-out mutants in Zea mays. New Phytol 218: 594–603 - PubMed
-
- Bosabalidis AM, Evert RF, Russin WA (1994) Ontogeny of the vascular bundles and contiguous tissues in the maize leaf blade. Am J Bot 81: 745–752
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
