

Pseudomonas aeruginosa-induced nociceptor activation increases susceptibility to infection
- PMID: 33956874
- PMCID: PMC8101935
- DOI: 10.1371/journal.ppat.1009557
Pseudomonas aeruginosa-induced nociceptor activation increases susceptibility to infection
Retraction in
-
Retraction: Pseudomonas aeruginosa-induced nociceptor activation increases susceptibility to infection.PLoS Pathog. 2024 Oct 18;20(10):e1012646. doi: 10.1371/journal.ppat.1012646. eCollection 2024 Oct. PLoS Pathog. 2024. PMID: 39423173 Free PMC article. No abstract available.
Abstract
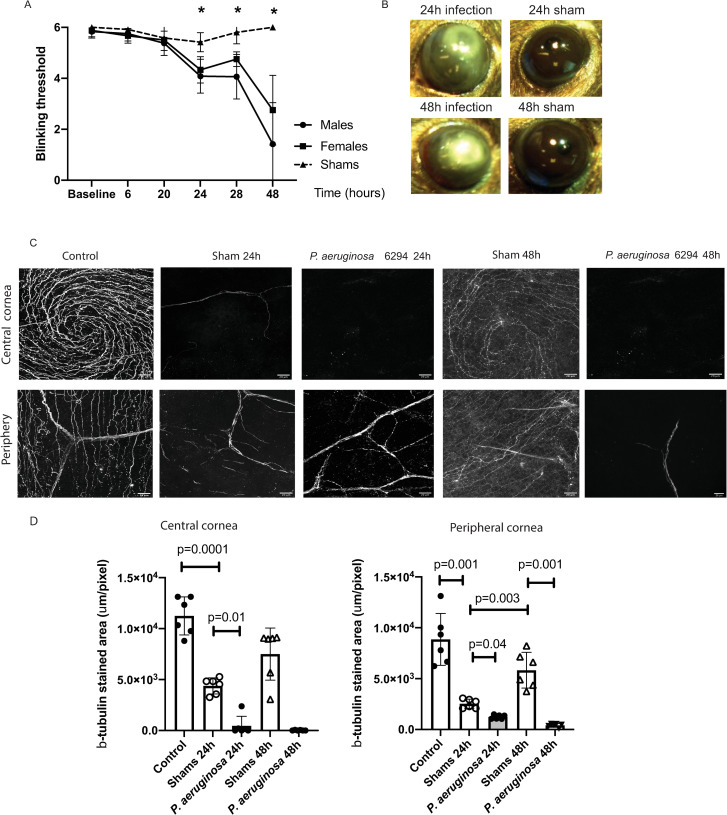
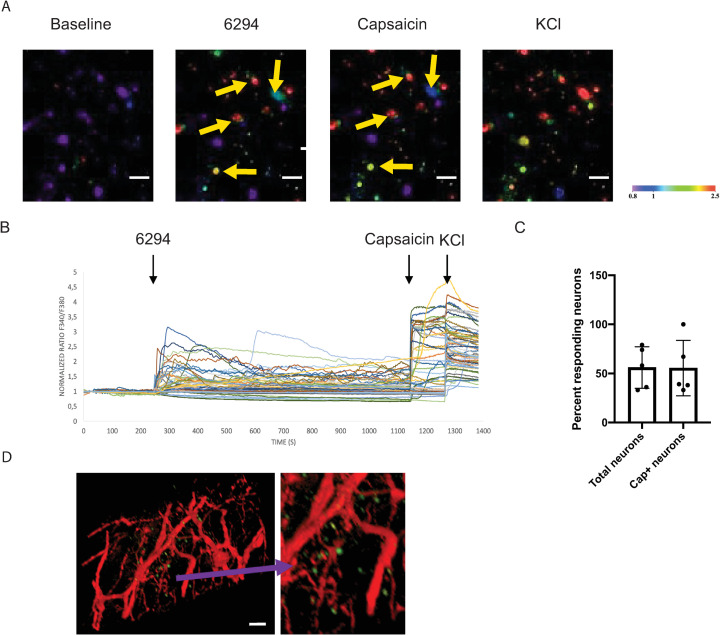
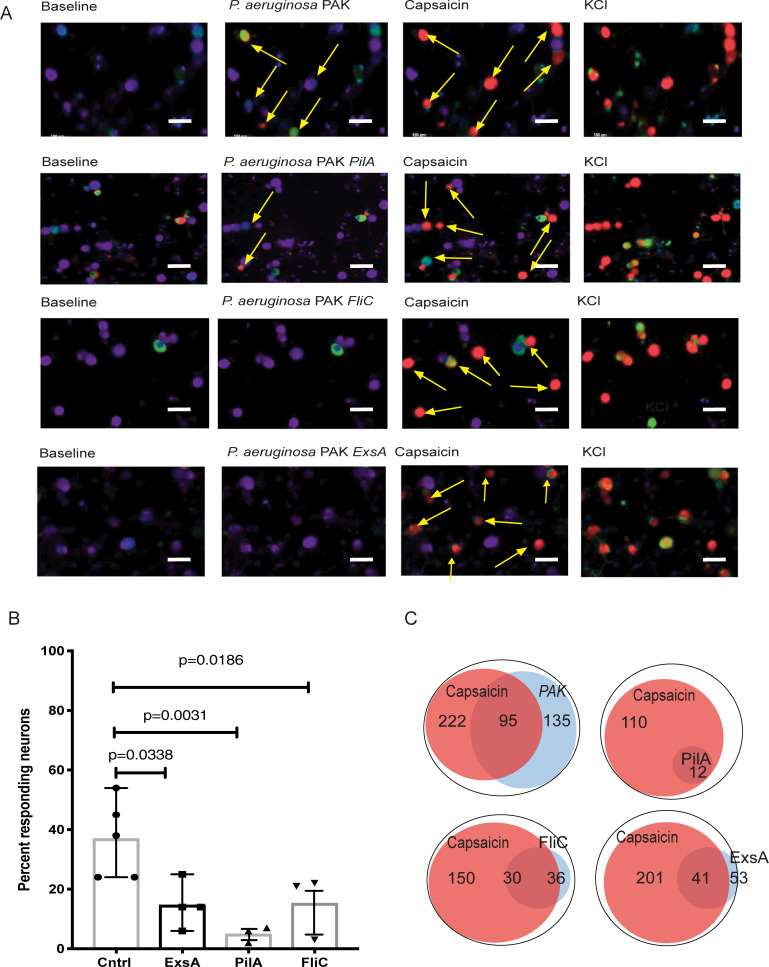
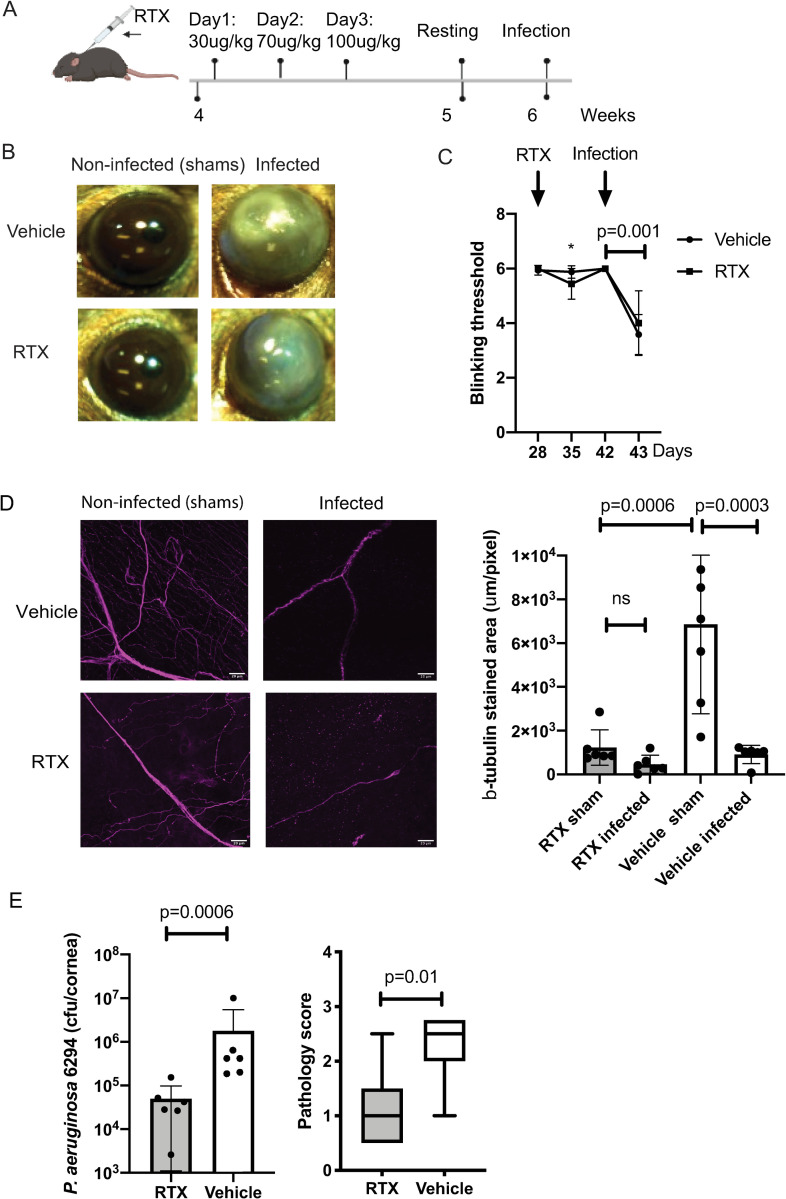
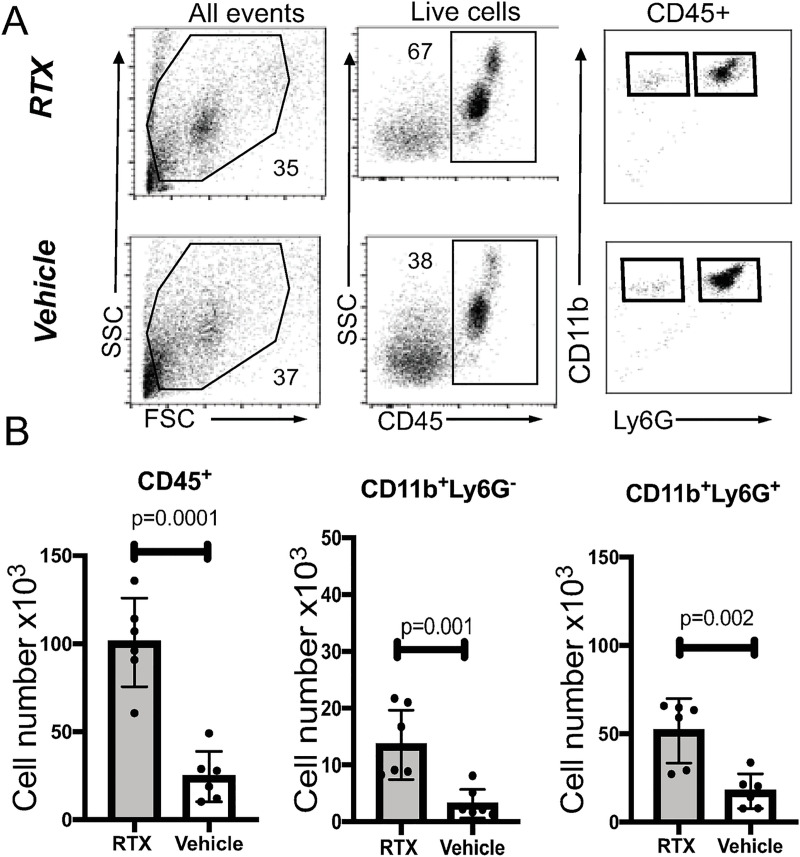
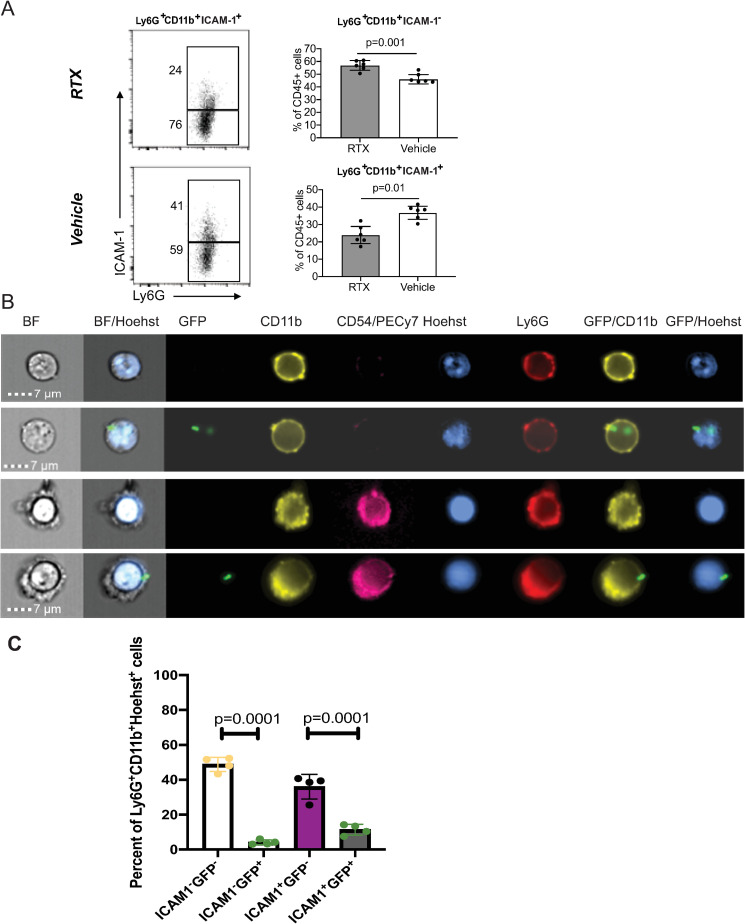
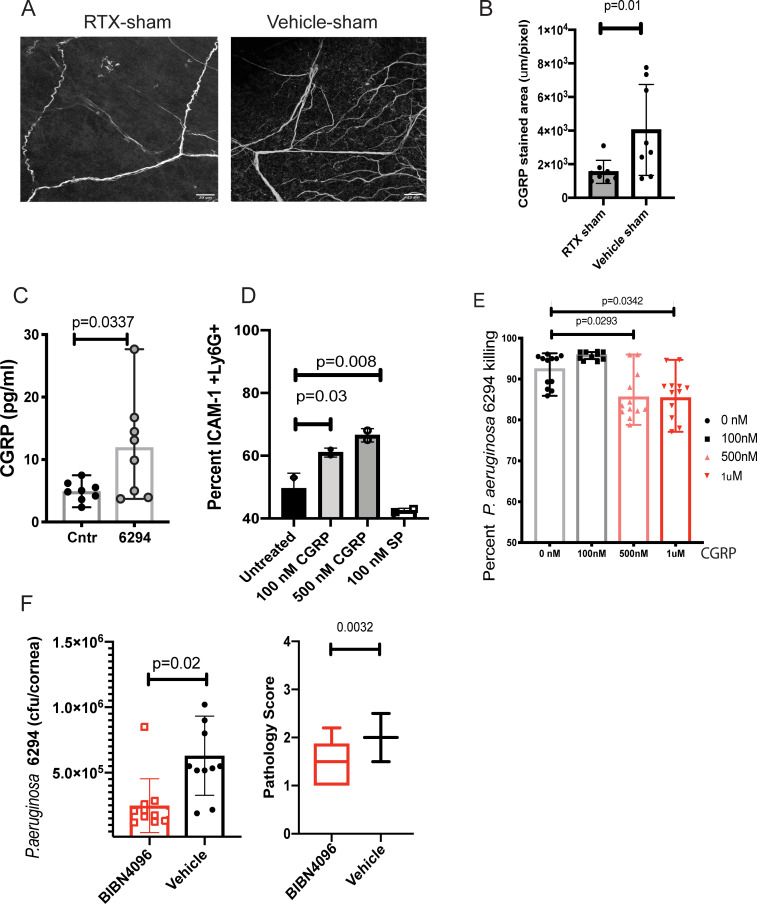
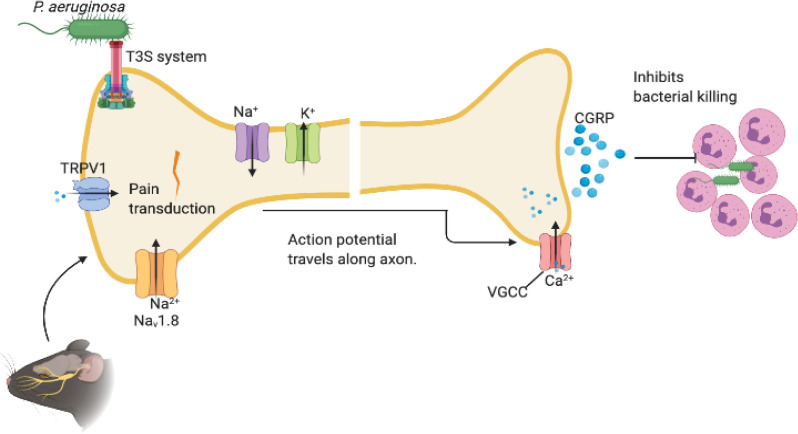
We report a rapid reduction in blink reflexes during in vivo ocular Pseudomonas aeruginosa infection, which is commonly attributed and indicative of functional neuronal damage. Sensory neurons derived in vitro from trigeminal ganglia (TG) were able to directly respond to P. aeruginosa but reacted significantly less to strains of P. aeruginosa that lacked virulence factors such as pili, flagella, or a type III secretion system. These observations led us to explore the impact of neurons on the host's susceptibility to P. aeruginosa keratitis. Mice were treated with Resiniferatoxin (RTX), a potent activator of Transient Receptor Potential Vanilloid 1 (TRPV1) channels, which significantly ablated corneal sensory neurons, exhibited delayed disease progression that was exemplified with decreased bacterial corneal burdens and altered neutrophil trafficking. Sensitization to disease was due to the increased frequencies of CGRP-induced ICAM-1+ neutrophils in the infected corneas and reduced neutrophil bactericidal activities. These data showed that sensory neurons regulate corneal neutrophil responses in a tissue-specific matter affecting disease progression during P. aeruginosa keratitis. Hence, therapeutic modalities that control nociception could beneficially impact anti-infective therapy.
Conflict of interest statement
No authors have competing interests.
Figures
References
-
- Weed MC RG, Kitzmann AS, Goins KM, Wagoner MD. V. Vision Loss After Contact Lens-Related Pseudomonas Keratitis. EyeRoundsorg. 2013.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
