

Structure and Mechanics of Dynein Motors
- PMID: 33957056
- PMCID: PMC8592121
- DOI: 10.1146/annurev-biophys-111020-101511
Structure and Mechanics of Dynein Motors
Abstract
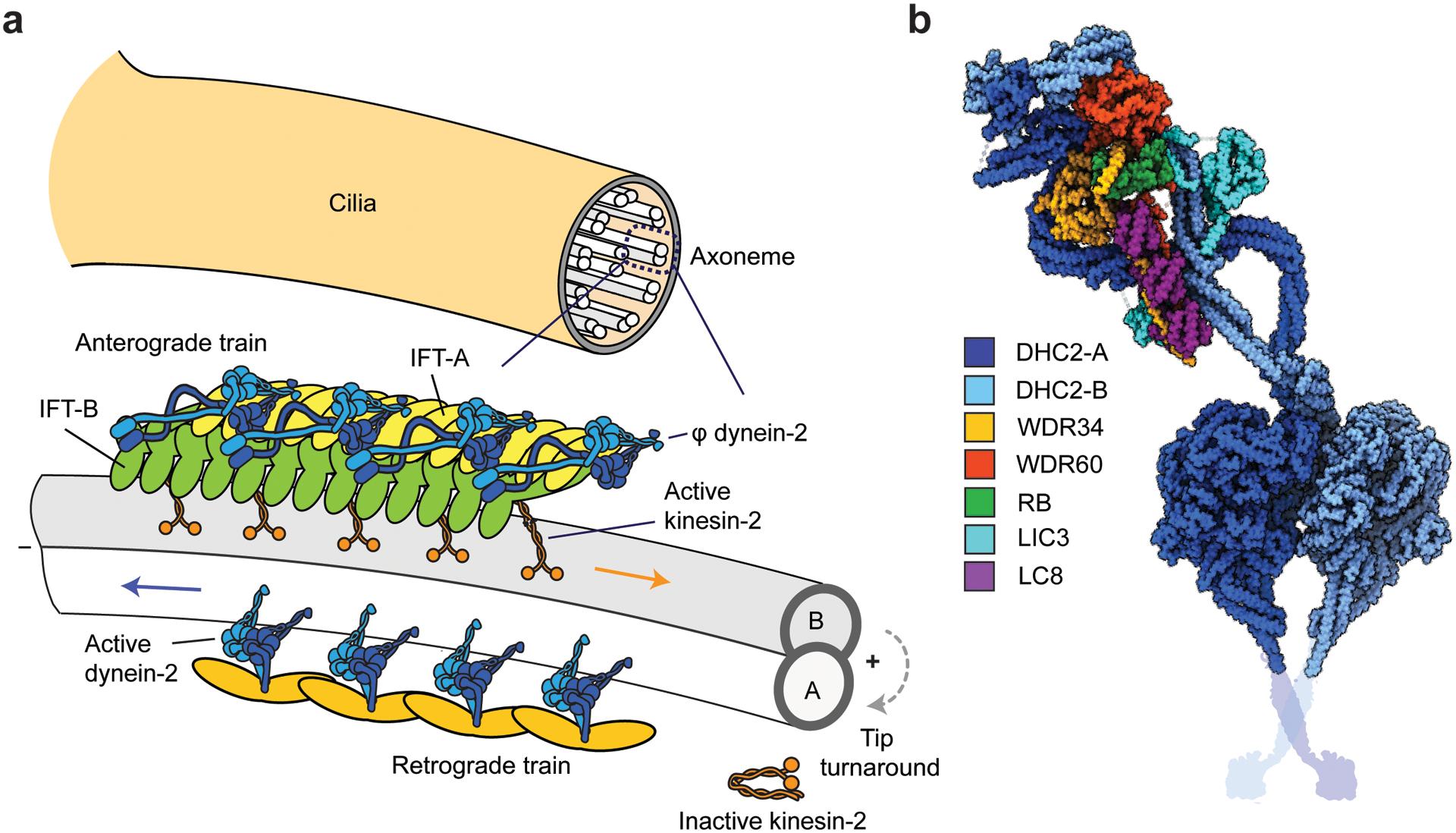
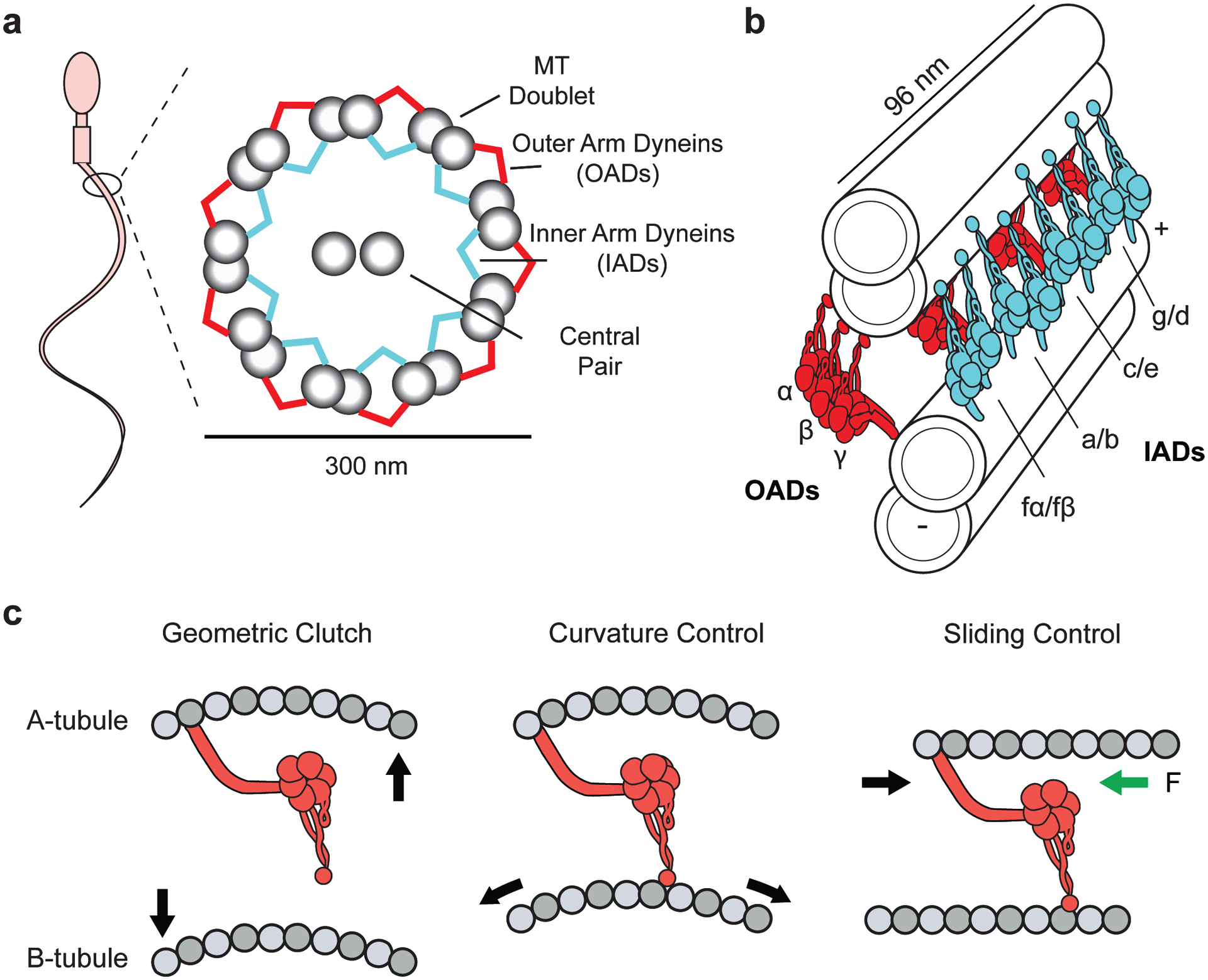
Dyneins make up a family of AAA+ motors that move toward the minus end of microtubules. Cytoplasmic dynein is responsible for transporting intracellular cargos in interphase cells and mediating spindle assembly and chromosome positioning during cell division. Other dynein isoforms transport cargos in cilia and power ciliary beating. Dyneins were the least studied of the cytoskeletal motors due to challenges in the reconstitution of active dynein complexes in vitro and the scarcity of high-resolution methods for in-depth structural and biophysical characterization of these motors. These challenges have been recently addressed, and there have been major advances in our understanding of the activation, mechanism, and regulation of dyneins. This review synthesizes the results of structural and biophysical studies for each class of dynein motors. We highlight several outstanding questions about the regulation of bidirectional transport along microtubules and the mechanisms that sustain self-coordinated oscillations within motile cilia.
Keywords: axoneme; cilia; dynein; intracellular transport; intraflagellar transport; microtubules.
Figures
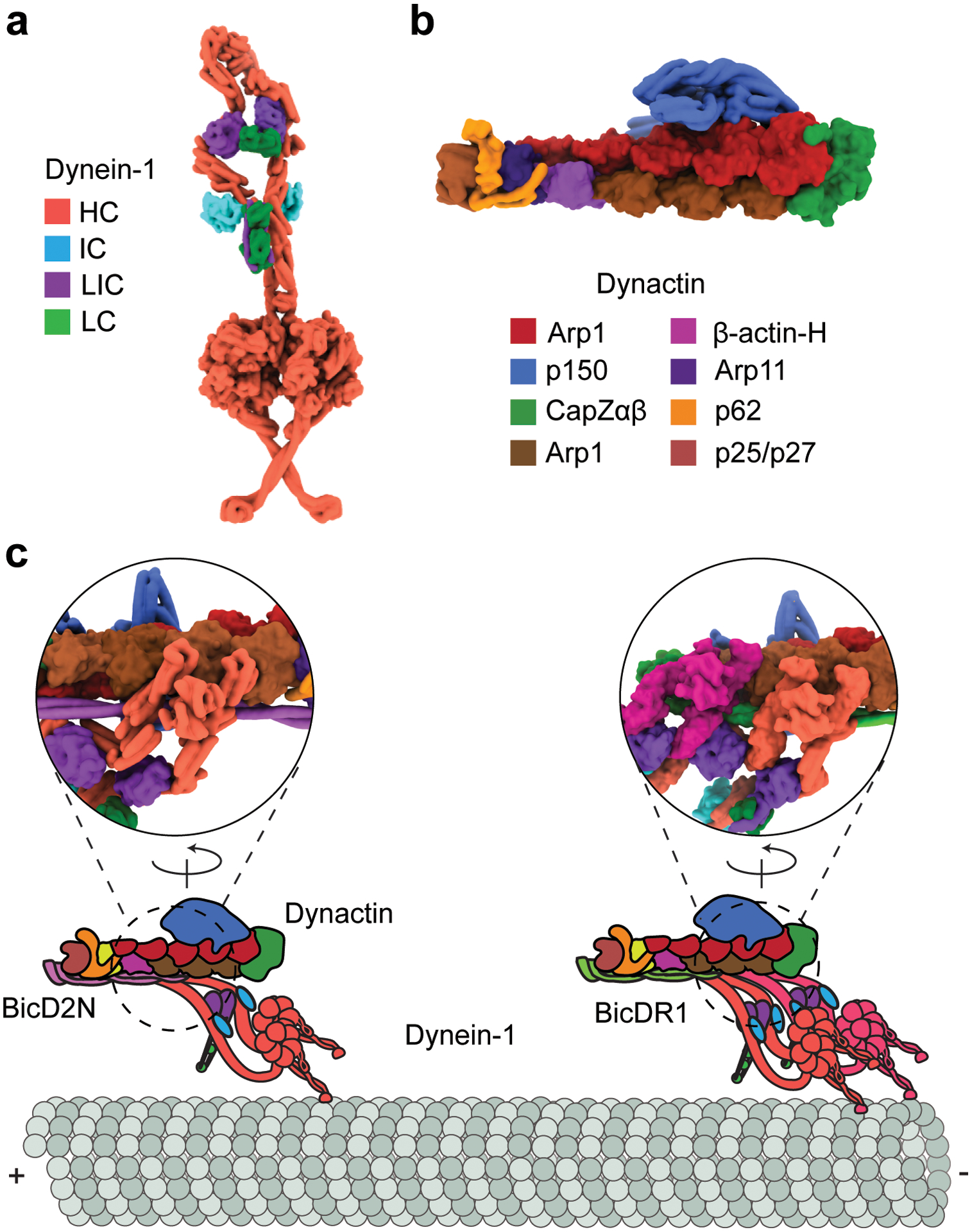
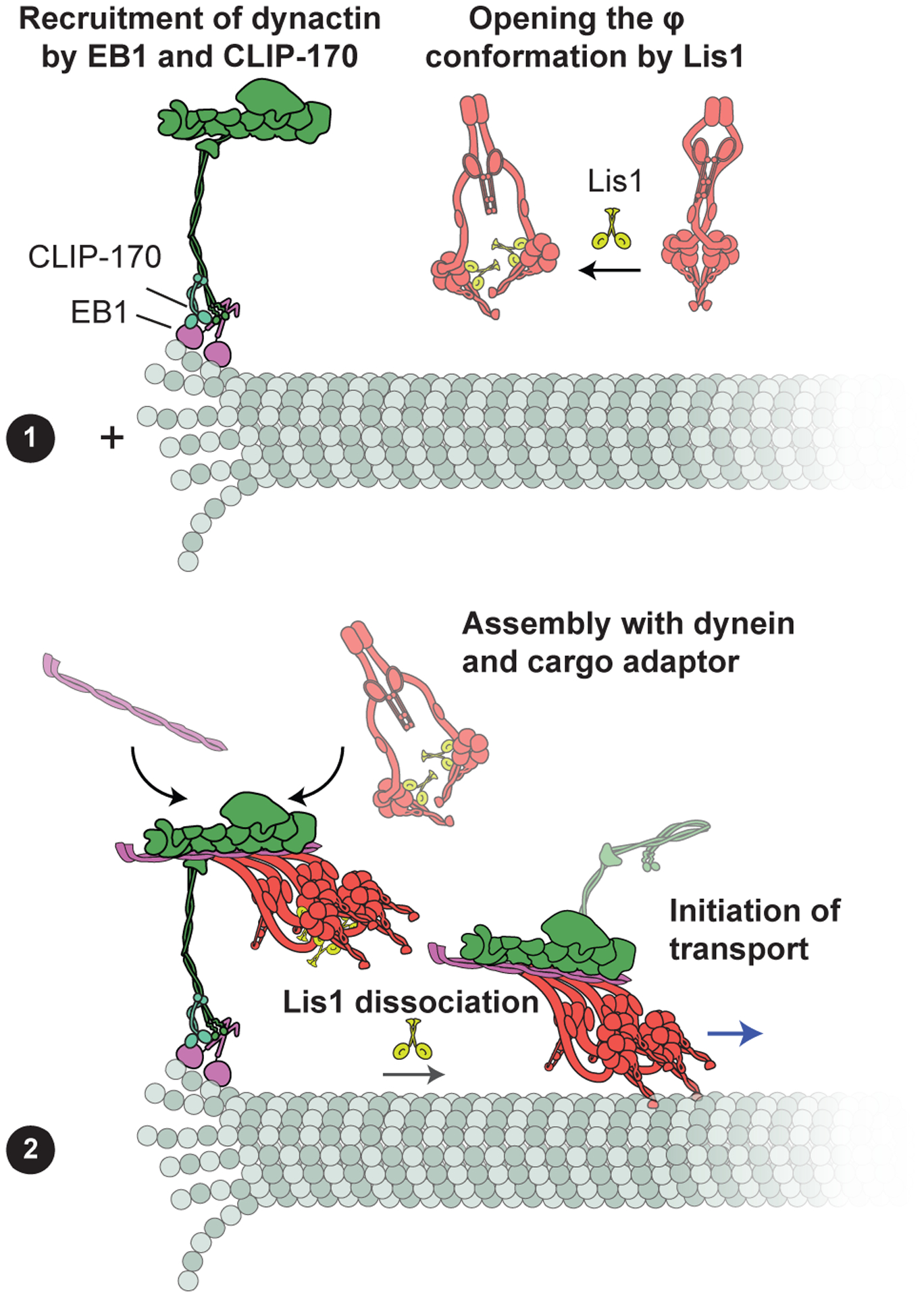
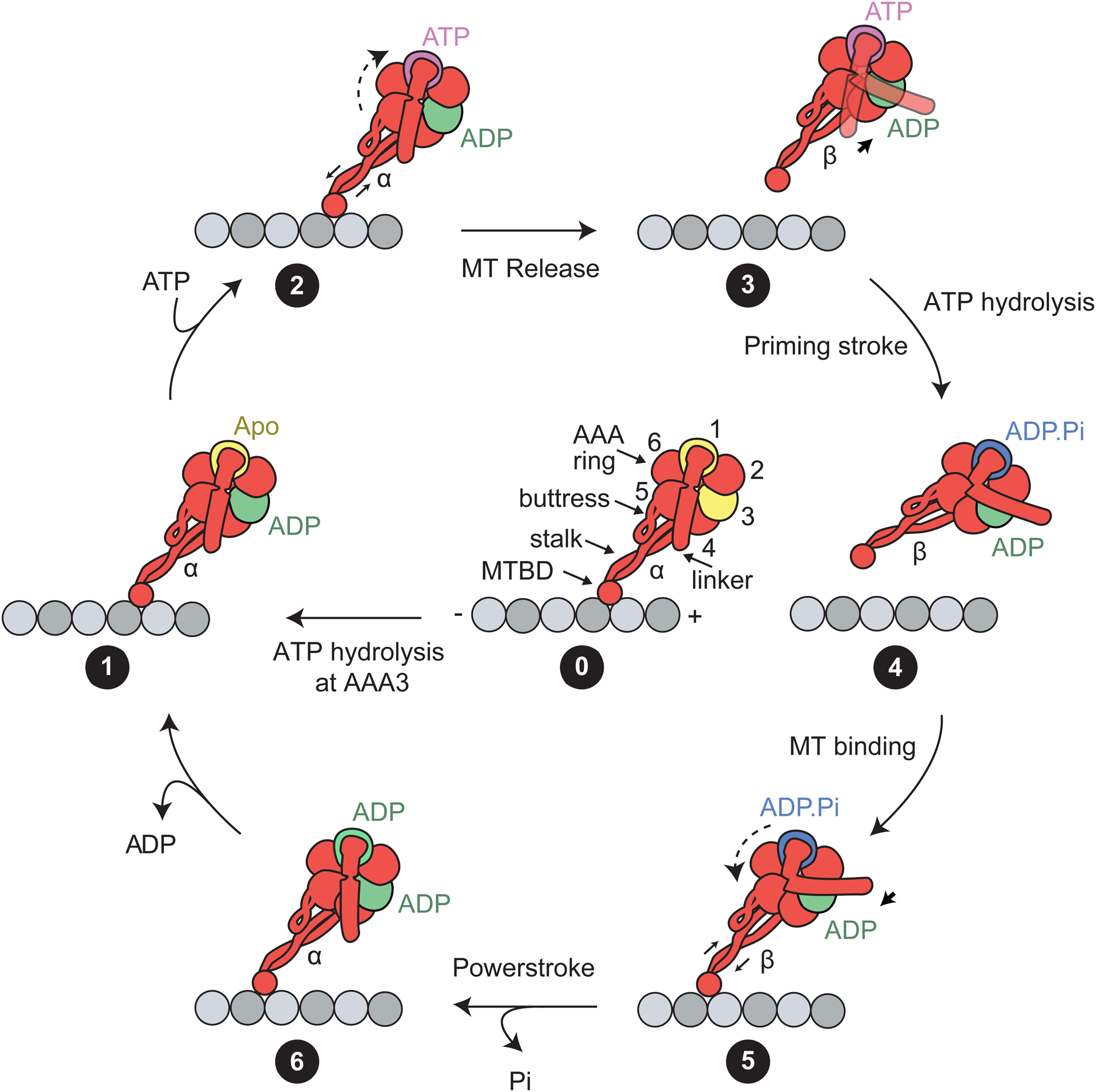
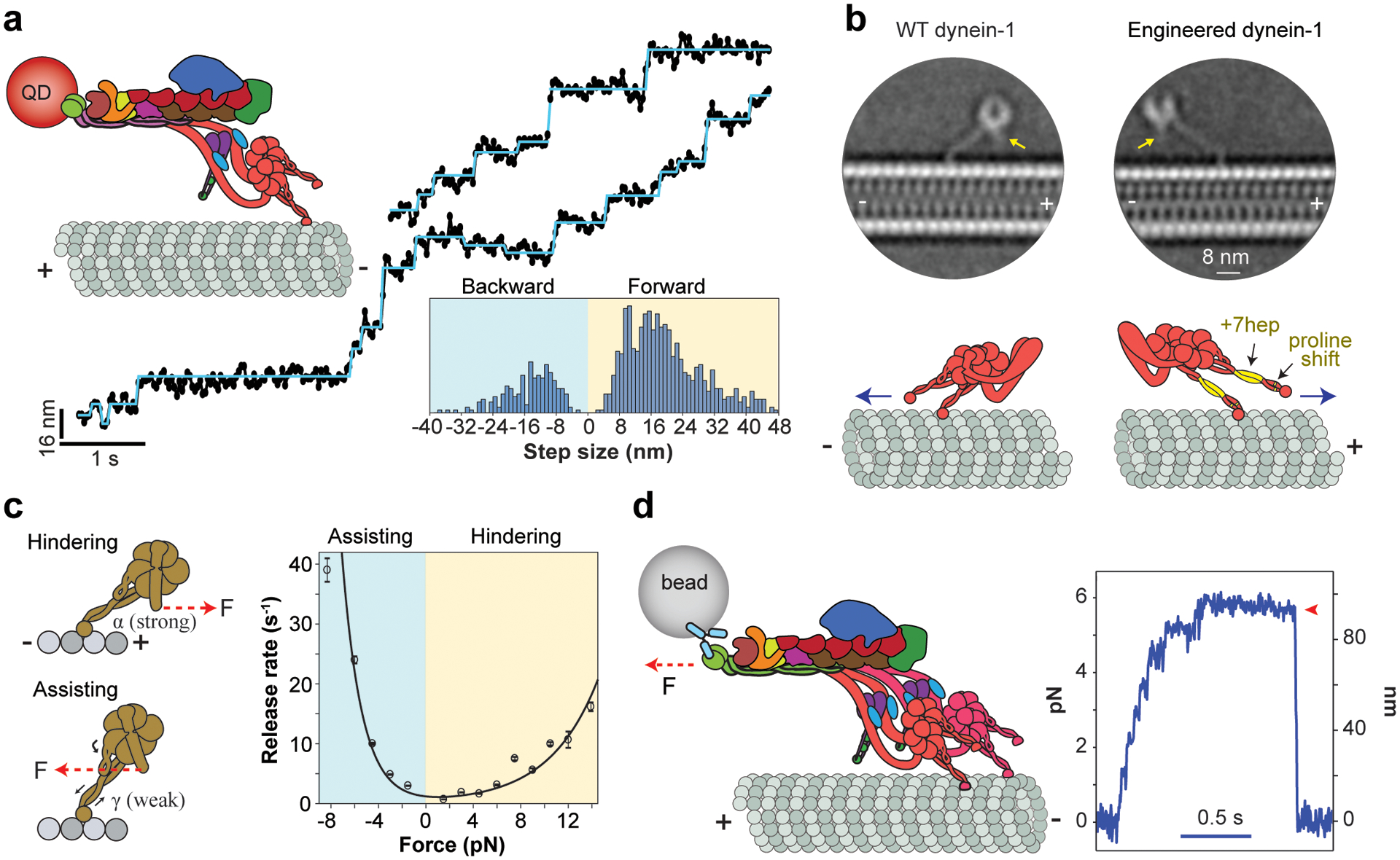
References
-
- Amos LA. 1989. Brain dynein crossbridges microtubules into bundles. J. Cell Sci 93(1):19–28 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
