

50 Years of structural immunology
- PMID: 33957119
- PMCID: PMC8163984
- DOI: 10.1016/j.jbc.2021.100745
50 Years of structural immunology
Abstract
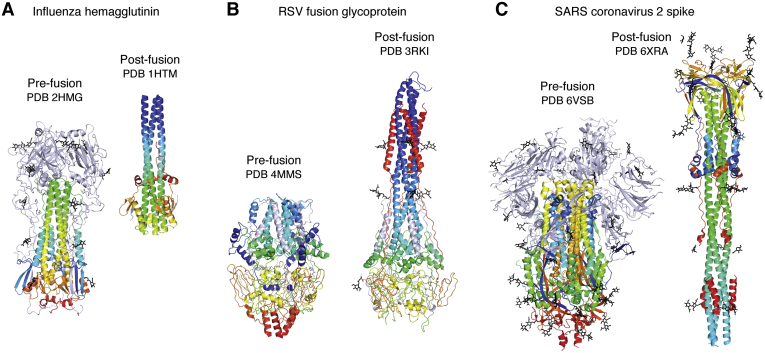
Fifty years ago, the first landmark structures of antibodies heralded the dawn of structural immunology. Momentum then started to build toward understanding how antibodies could recognize the vast universe of potential antigens and how antibody-combining sites could be tailored to engage antigens with high specificity and affinity through recombination of germline genes (V, D, J) and somatic mutation. Equivalent groundbreaking structures in the cellular immune system appeared some 15 to 20 years later and illustrated how processed protein antigens in the form of peptides are presented by MHC molecules to T cell receptors. Structures of antigen receptors in the innate immune system then explained their inherent specificity for particular microbial antigens including lipids, carbohydrates, nucleic acids, small molecules, and specific proteins. These two sides of the immune system act immediately (innate) to particular microbial antigens or evolve (adaptive) to attain high specificity and affinity to a much wider range of antigens. We also include examples of other key receptors in the immune system (cytokine receptors) that regulate immunity and inflammation. Furthermore, these antigen receptors use a limited set of protein folds to accomplish their various immunological roles. The other main players are the antigens themselves. We focus on surface glycoproteins in enveloped viruses including SARS-CoV-2 that enable entry and egress into host cells and are targets for the antibody response. This review covers what we have learned over the past half century about the structural basis of the immune response to microbial pathogens and how that information can be utilized to design vaccines and therapeutics.
Keywords: MHC; T cells; TLR; VLR; antibodies; cellular immunity; humoral immunity; immune recognition; microbial pathogens; viral antigens.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
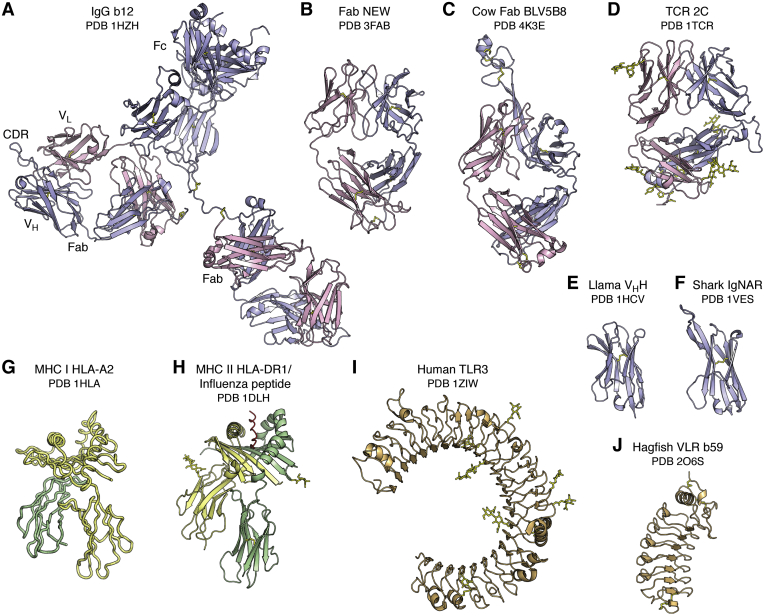
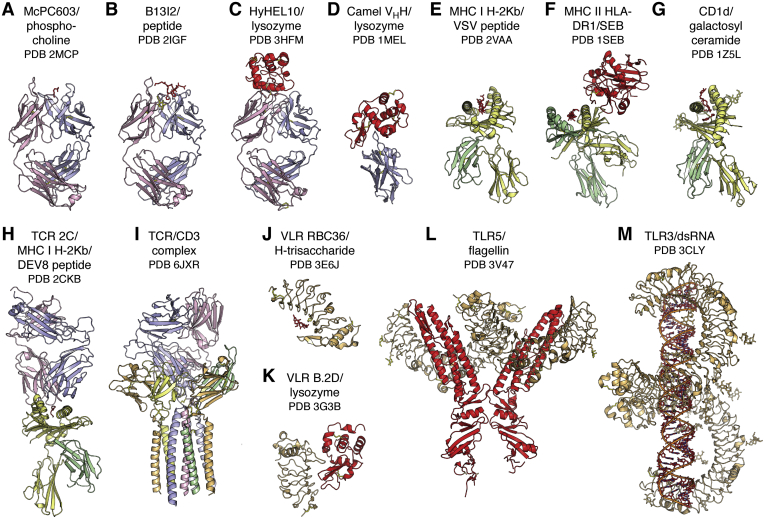
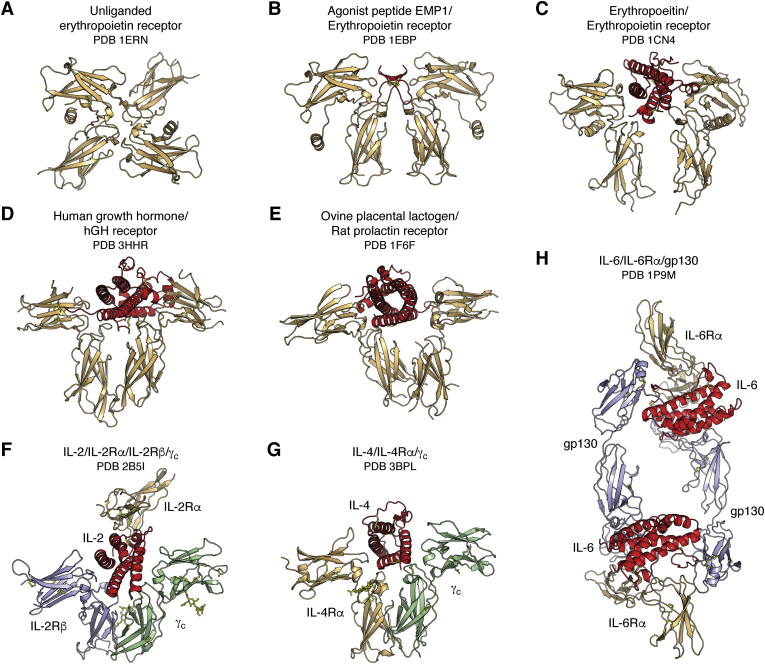
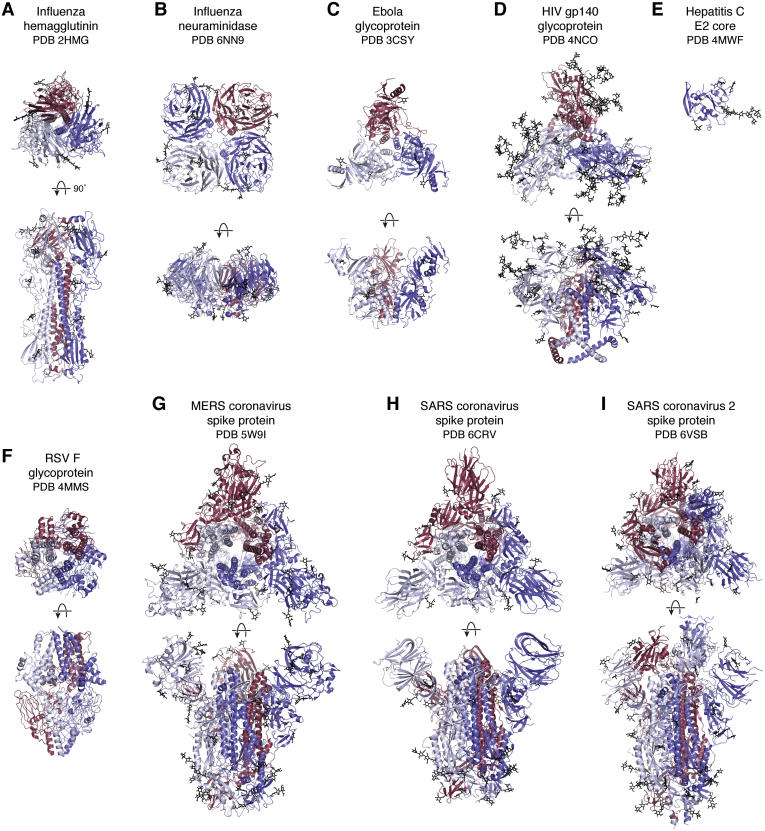

References
-
- Sarma V.R., Silverton E.W., Davies D.R., Terry W.D. The three-dimensional structure at 6 Å resolution of a human γG1 immunoglobulin molecule. J. Biol. Chem. 1971;246:3753–3759. - PubMed
-
- Poljak R.J., Amzel L.M., Avey H.P., Becka L.N. Structure of Fab’ New at 6 Å resolution. Nat. New Biol. 1972;235:137–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
