

Genetic and environmental modulation of transposition shapes the evolutionary potential of Arabidopsis thaliana
- PMID: 33957946
- PMCID: PMC8101250
- DOI: 10.1186/s13059-021-02348-5
Genetic and environmental modulation of transposition shapes the evolutionary potential of Arabidopsis thaliana
Abstract
Background: How species can adapt to abrupt environmental changes, particularly in the absence of standing genetic variation, is poorly understood and a pressing question in the face of ongoing climate change. Here we leverage publicly available multi-omic and bio-climatic data for more than 1000 wild Arabidopsis thaliana accessions to determine the rate of transposable element (TE) mobilization and its potential to create adaptive variation in natural settings.
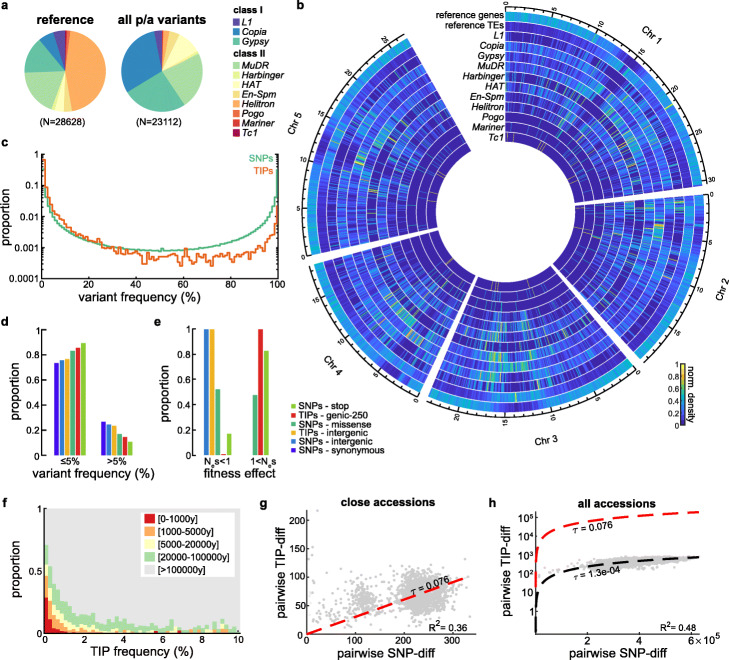
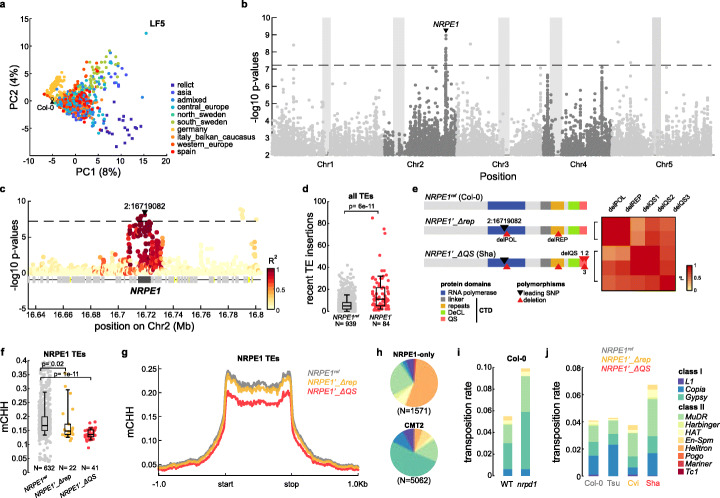
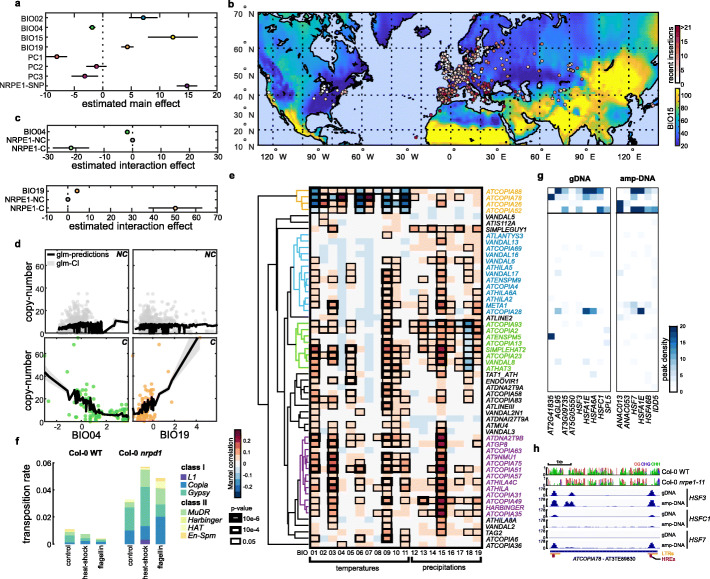
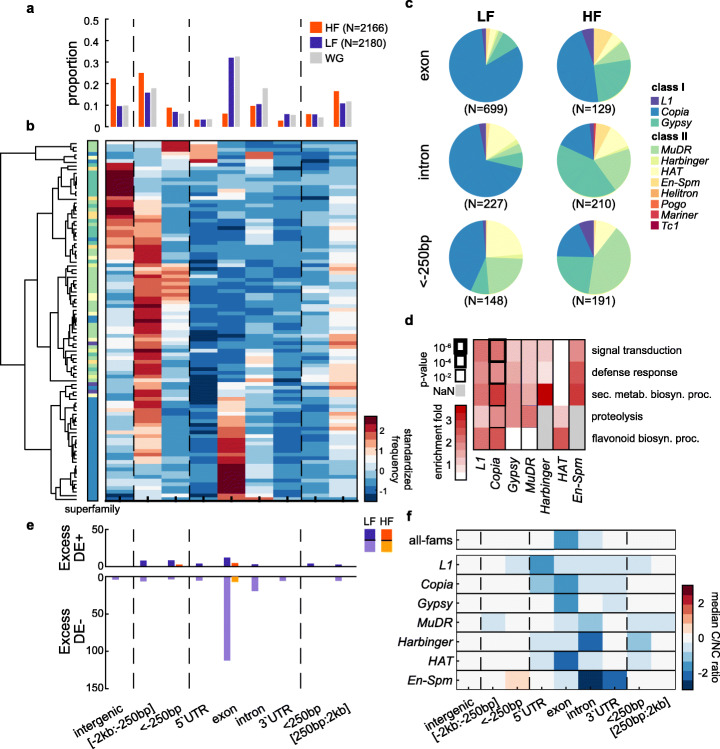
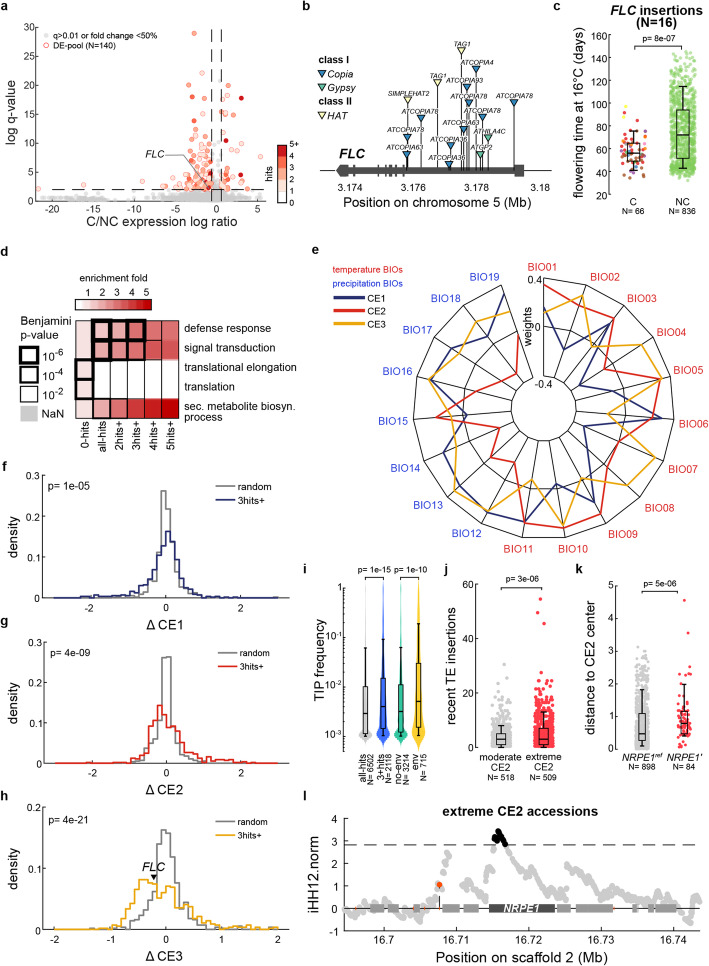
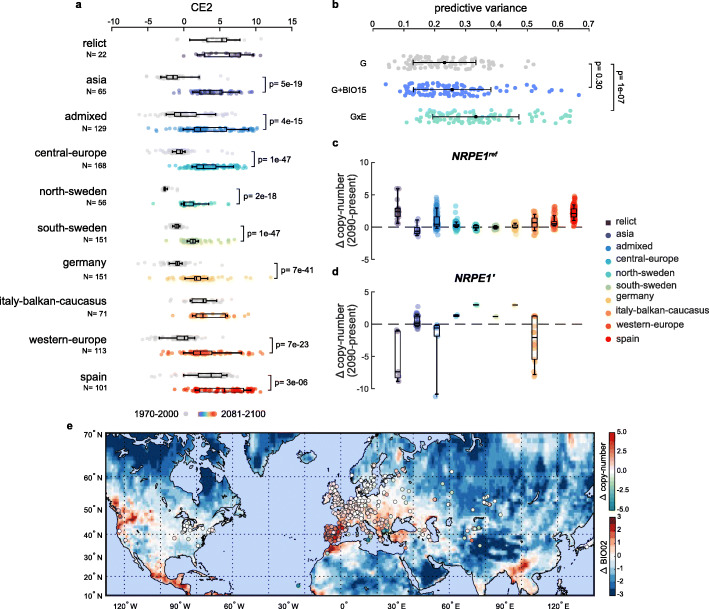
Results: We demonstrate that TE insertions arise at almost the same rate as base substitutions. Mobilization activity of individual TE families varies greatly between accessions, in association with genetic and environmental factors as well as through complex gene-environment interactions. Although the distribution of TE insertions across the genome is ultimately shaped by purifying selection, reflecting their typically strong deleterious effects when located near or within genes, numerous recent TE-containing alleles show signatures of positive selection. Moreover, high rates of transposition appear positively selected at the edge of the species' ecological niche. Based on these findings, we predict through mathematical modeling higher transposition activity in Mediterranean regions within the next decades in response to global warming, which in turn should accelerate the creation of large-effect alleles.
Conclusions: Our study reveals that TE mobilization is a major generator of genetic variation in A. thaliana that is finely modulated by genetic and environmental factors. These findings and modeling indicate that TEs may be essential genomic players in the demise or rescue of native populations in times of climate crises.
Keywords: Adaptation; Climate change; Epigenomics; Genome evolution; Population genetics; Transposable elements.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Barrett R, Schluter D. Adaptation from standing genetic variation. Trends Ecol Evol. 2008:38–44. 10.1016/j.tree.2007.09.008 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
