

Endothelial angiogenic activity and adipose angiogenesis is controlled by extracellular matrix protein TGFBI
- PMID: 33958649
- PMCID: PMC8102489
- DOI: 10.1038/s41598-021-88959-1
Endothelial angiogenic activity and adipose angiogenesis is controlled by extracellular matrix protein TGFBI
Abstract
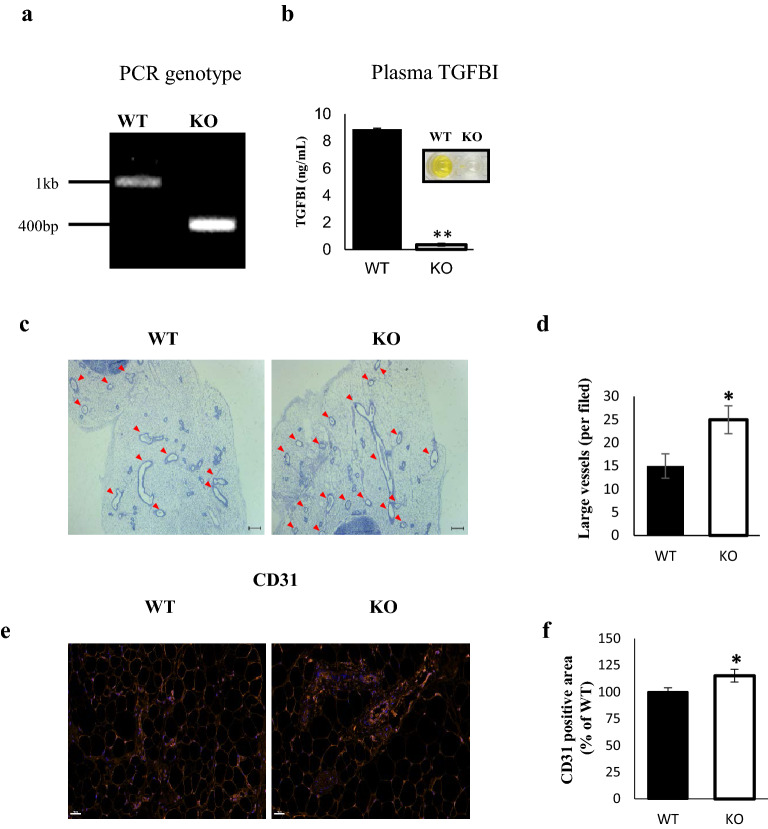
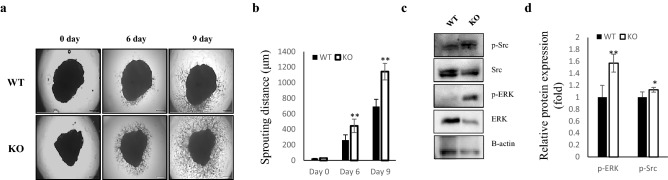
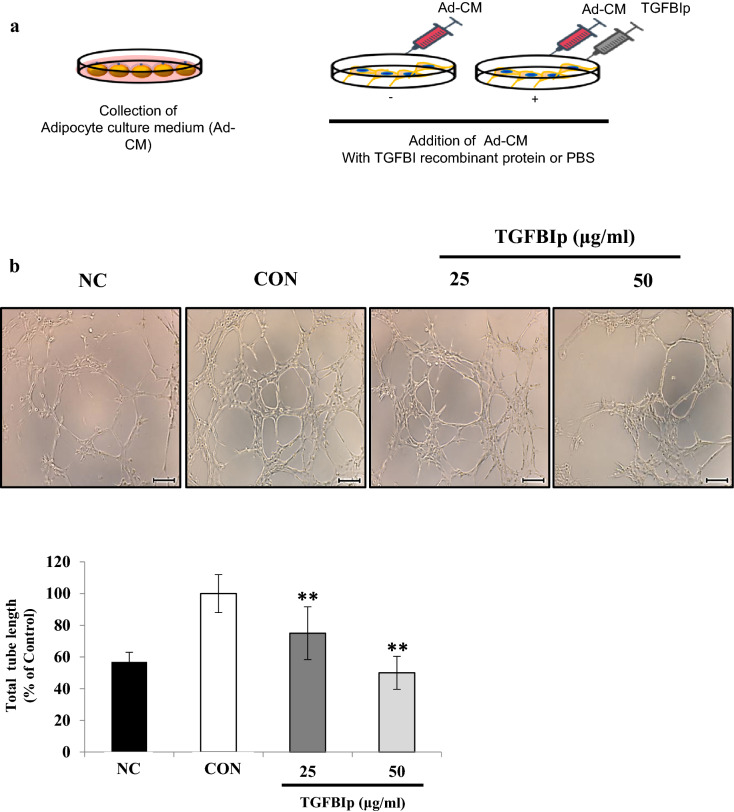
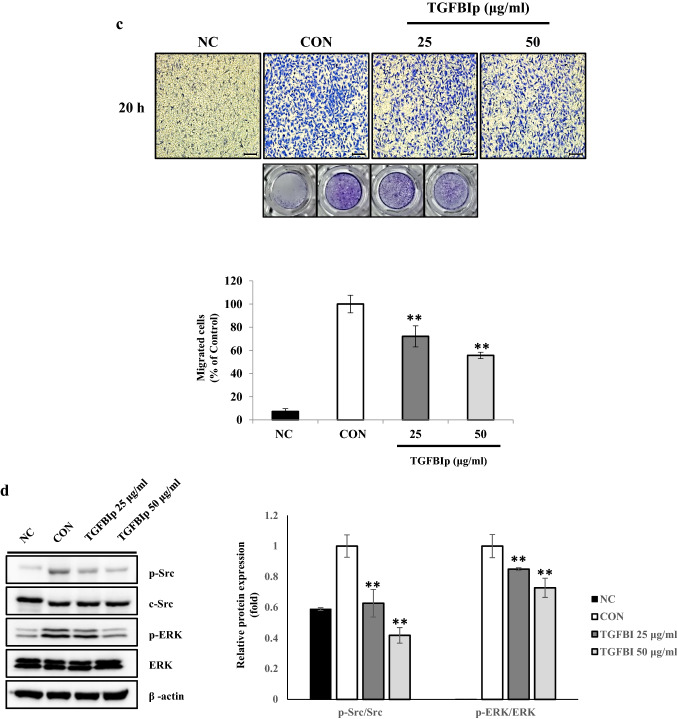
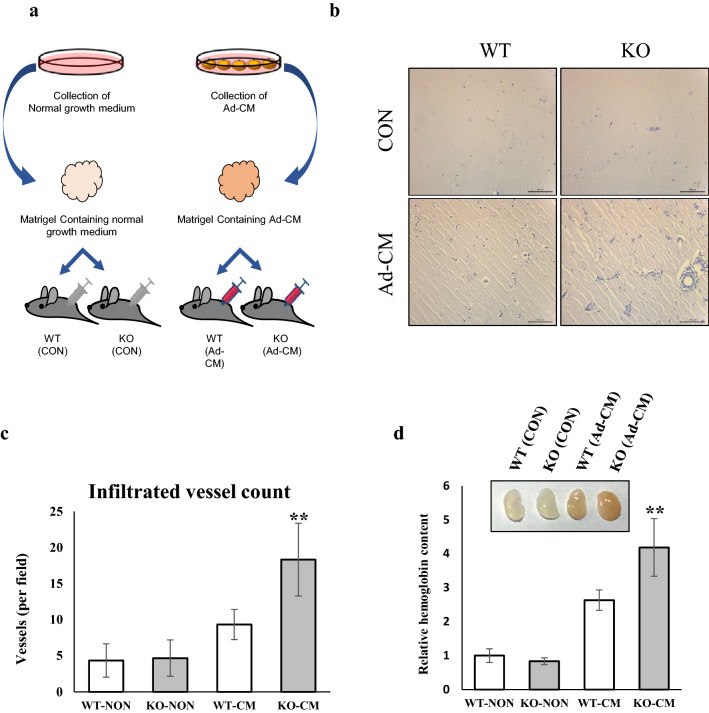
Several studies have suggested that extracellular matrix (ECM) remodeling and the microenvironment are tightly associated with adipogenesis and adipose angiogenesis. In the present study, we demonstrated that transforming growth factor-beta induced (TGFBI) suppresses angiogenesis stimulated by adipocyte-conditioned medium (Ad-CM), both in vitro and in vivo. TGFBI knockout (KO) mice exhibited increased numbers of blood vessels in adipose tissue, and blood vessels from these mice showed enhanced infiltration into Matrigel containing Ad-CM. The treatment of Ad-CM-stimulated SVEC-10 endothelial cells with TGFBI protein reduced migration and tube-forming activity. TGFBI protein suppressed the activation of the Src and extracellular signaling-related kinase signaling pathways of these SVEC-10 endothelial cells. Our findings indicated that TGFBI inhibited adipose angiogenesis by suppressing the activation of Src and ERK signaling pathways, possibly because of the stimulation of the angiogenic activity of endothelial cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
