

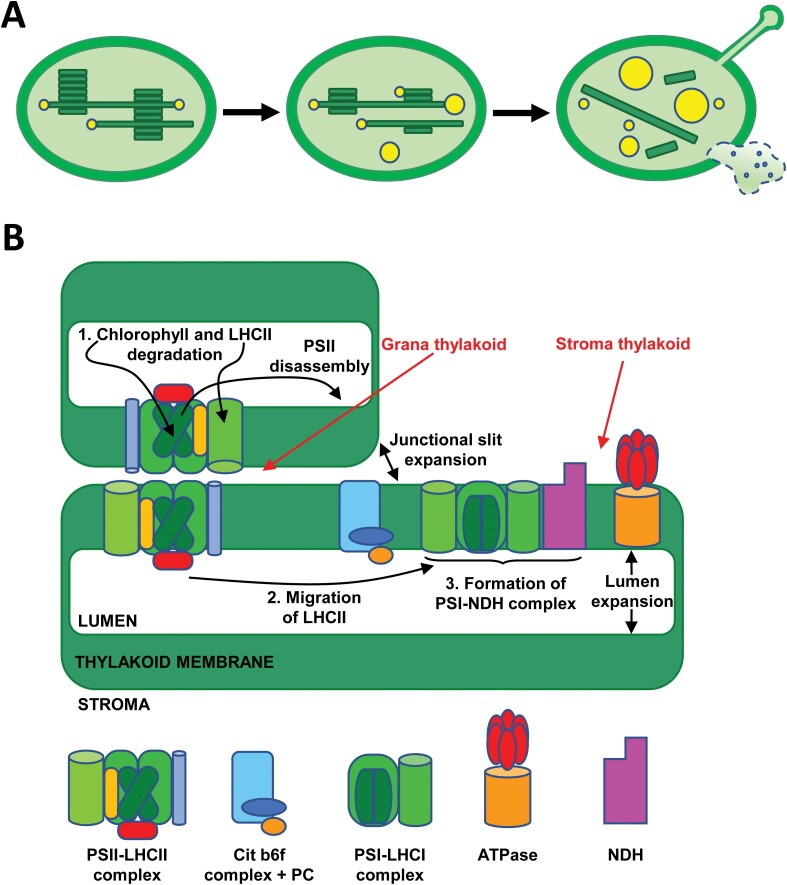
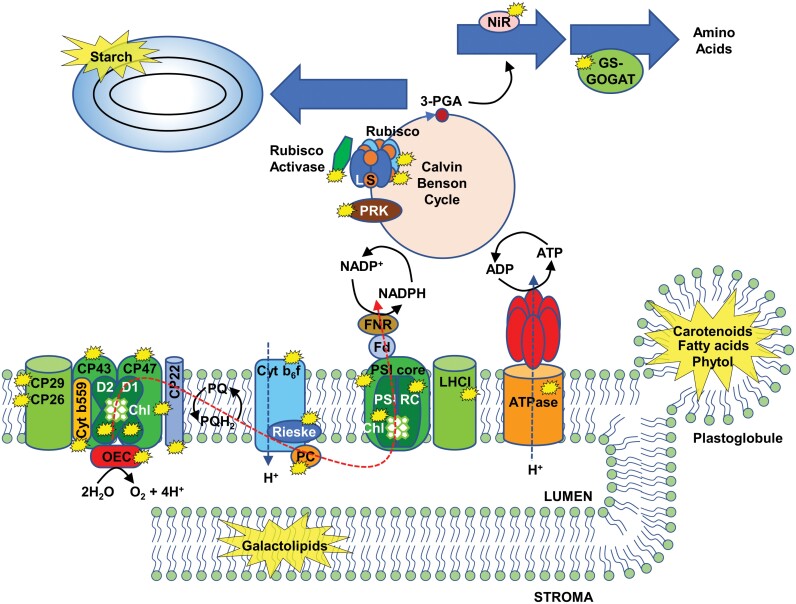
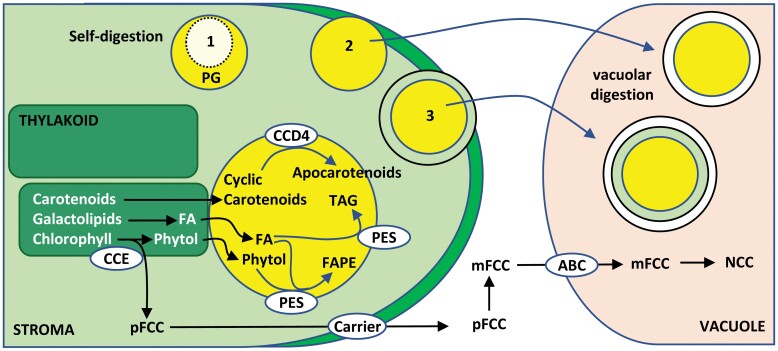
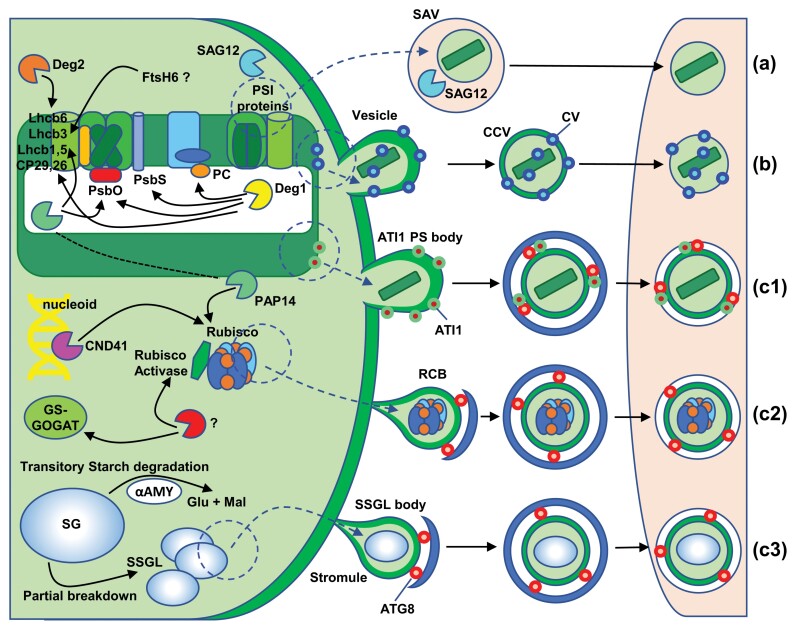
Chloroplast dismantling in leaf senescence
- PMID: 33959761
- PMCID: PMC8760853
- DOI: 10.1093/jxb/erab200
Chloroplast dismantling in leaf senescence
Abstract
In photosynthetic plant cells, chloroplasts act as factories of metabolic intermediates that support plant growth. Chloroplast performance is highly influenced by environmental cues. Thus, these organelles have the additional function of sensing ever changing environmental conditions, thereby playing a key role in harmonizing the growth and development of different organs and in plant acclimation to the environment. Moreover, chloroplasts constitute an excellent source of metabolic intermediates that are remobilized to sink tissues during senescence so that chloroplast dismantling is a tightly regulated process that plays a key role in plant development. Stressful environmental conditions enhance the generation of reactive oxygen species (ROS) by chloroplasts, which may lead to oxidative stress causing damage to the organelle. These environmental conditions trigger mechanisms that allow the rapid dismantling of damaged chloroplasts, which is crucial to avoid deleterious effects of toxic by-products of the degradative process. In this review, we discuss the effect of redox homeostasis and ROS generation in the process of chloroplast dismantling. Furthermore, we summarize the structural and biochemical events, both intra- and extraplastid, that characterize the process of chloroplast dismantling in senescence and in response to environmental stresses.
Keywords: Autophagy; ROS; chlorophagy; chloroplast; gerontoplast; plastoglobule; senescence.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Avila-Ospina L, Moison M, Yoshimoto K, Masclaux-Daubresse C. 2014. Autophagy, plant senescence, and nutrient recycling. Journal of Experimental Botany 65, 3799–3811. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
