

Aldoximes are precursors of auxins in Arabidopsis and maize
- PMID: 33959967
- PMCID: PMC8282758
- DOI: 10.1111/nph.17447
Aldoximes are precursors of auxins in Arabidopsis and maize
Abstract
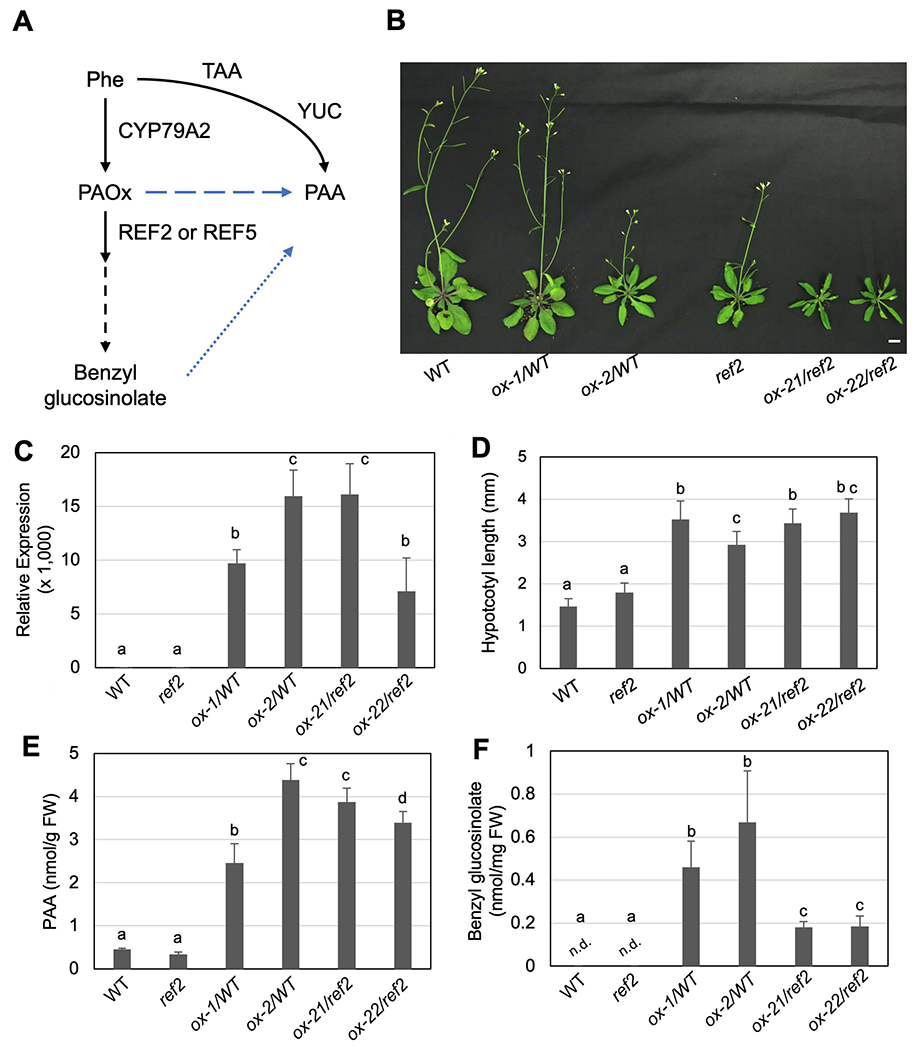
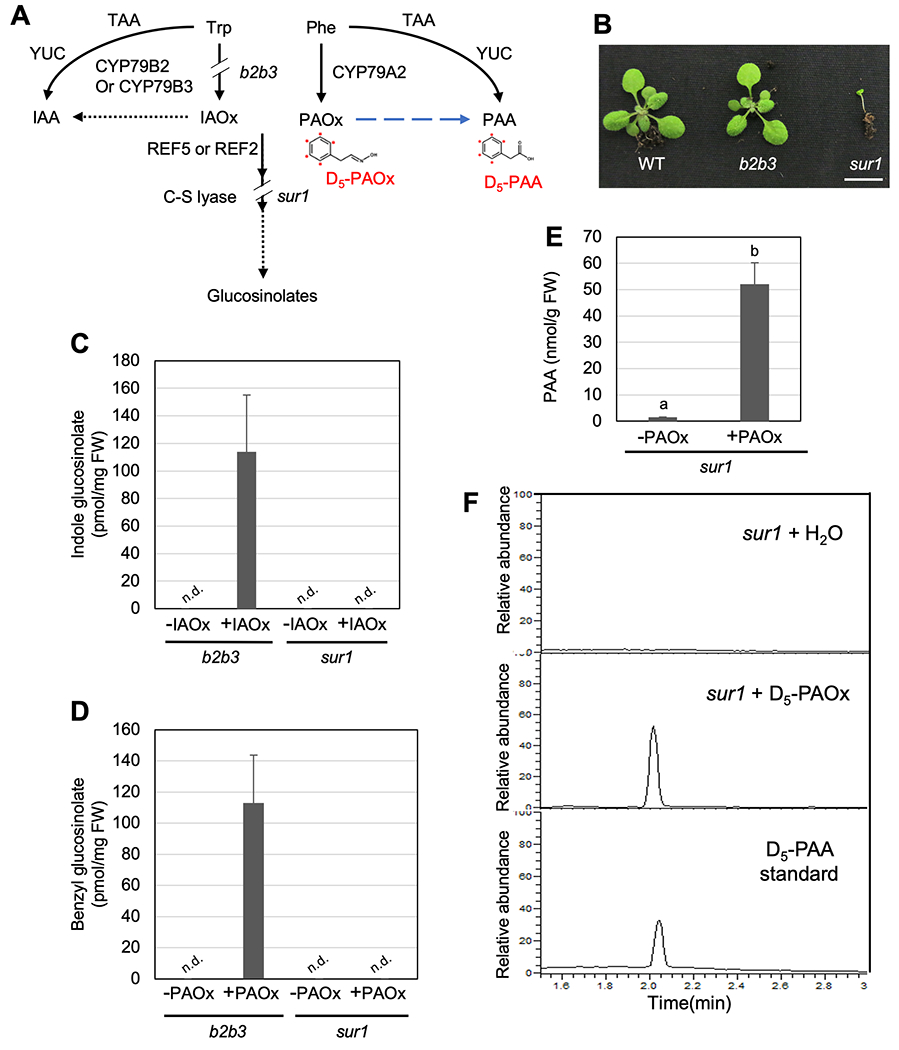
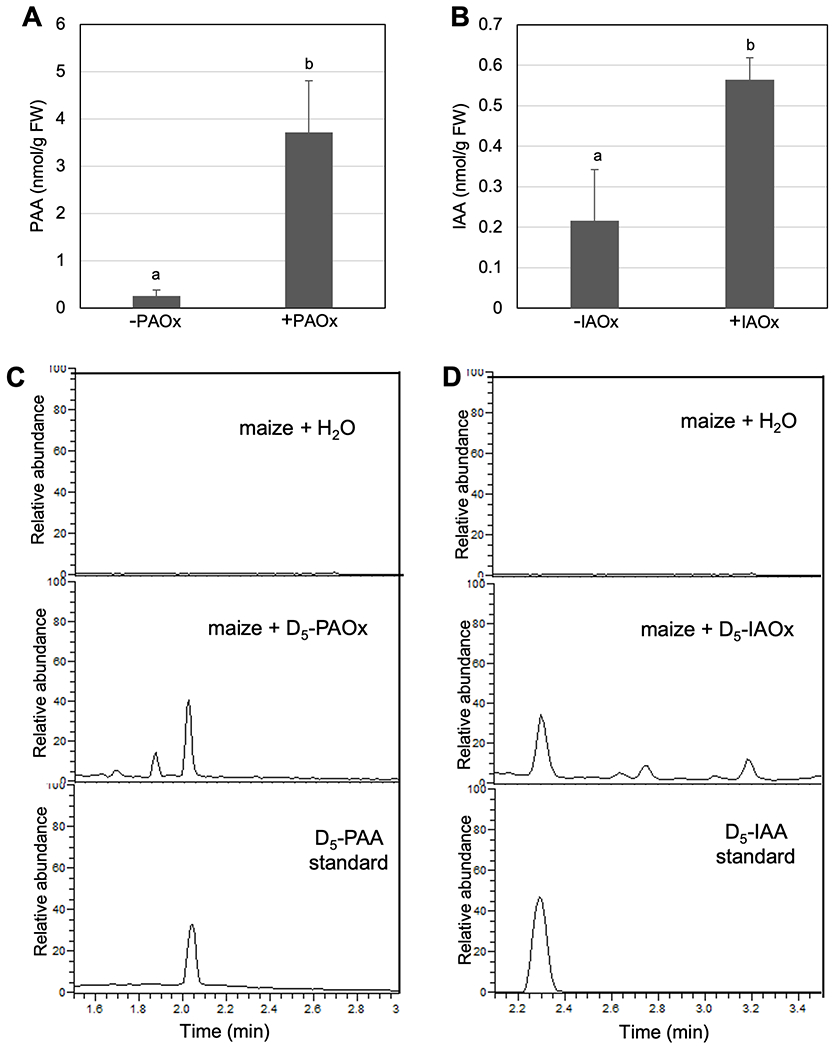
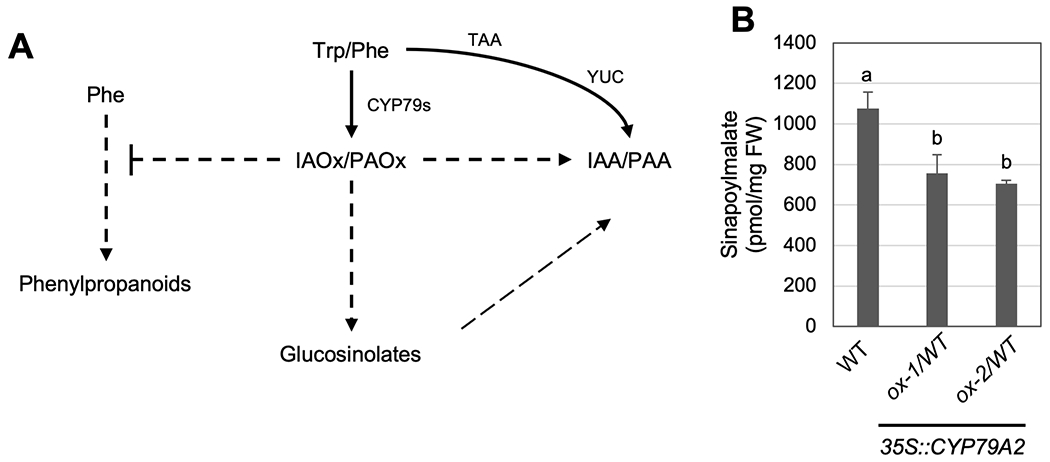
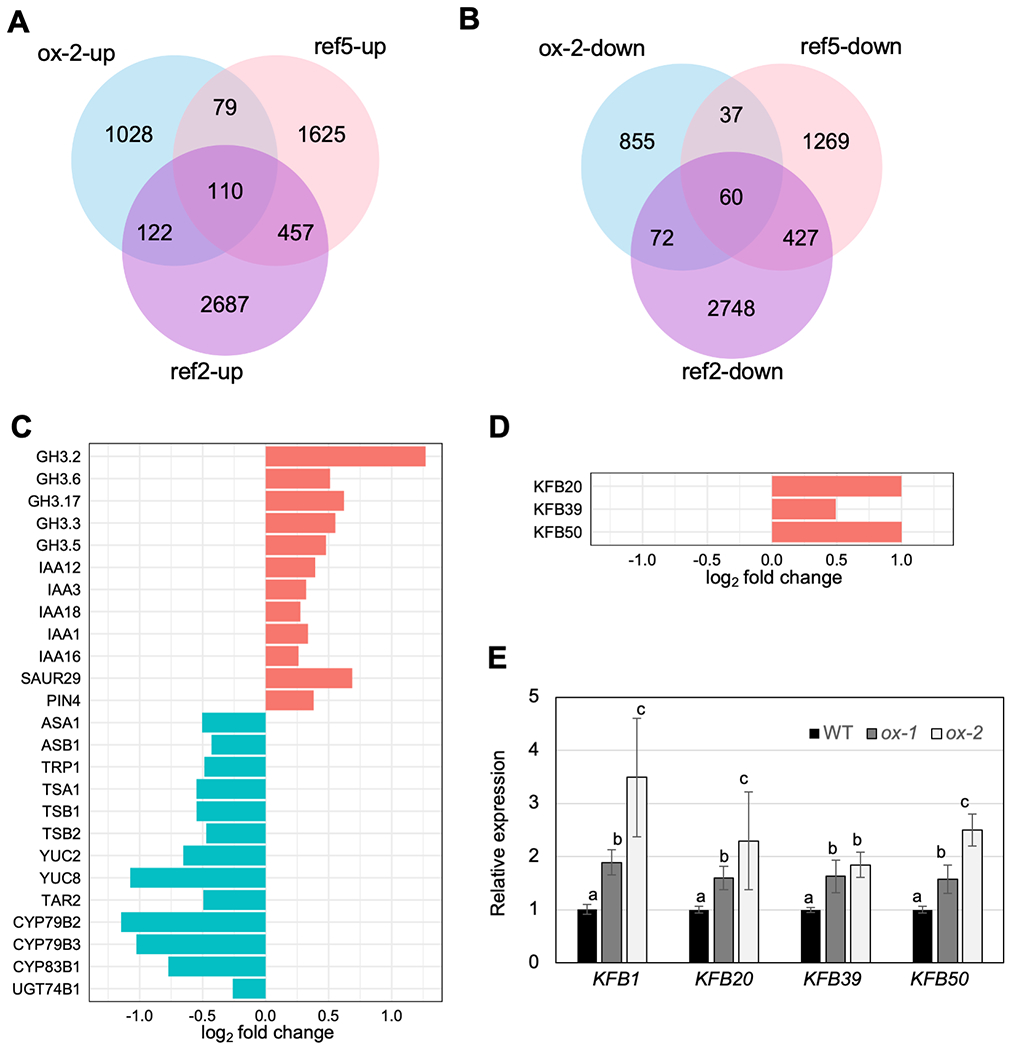
Two natural auxins, phenylacetic acid (PAA) and indole-3-acetic acid (IAA), play crucial roles in plant growth and development. One route of IAA biosynthesis uses the glucosinolate intermediate indole-3-acetaldoxime (IAOx) as a precursor, which is thought to occur only in glucosinolate-producing plants in Brassicales. A recent study showed that overproducing phenylacetaldoxime (PAOx) in Arabidopsis increases PAA production. However, it remains unknown whether this increased PAA resulted from hydrolysis of PAOx-derived benzyl glucosinolate or, like IAOx-derived IAA, is directly converted from PAOx. If glucosinolate hydrolysis is not required, aldoxime-derived auxin biosynthesis may occur beyond Brassicales. To better understand aldoxime-derived auxin biosynthesis, we conducted an isotope-labelled aldoxime feeding assay using an Arabidopsis glucosinolate-deficient mutant sur1 and maize, and transcriptomics analysis. Our study demonstrated that the conversion of PAOx to PAA does not require glucosinolates in Arabidopsis. Furthermore, maize produces PAA and IAA from PAOx and IAOx, respectively, indicating that aldoxime-derived auxin biosynthesis also occurs in maize. Considering that aldoxime production occurs widely in the plant kingdom, aldoxime-derived auxin biosynthesis is likely to be more widespread than originally believed. A genome-wide transcriptomics study using PAOx-overproduction plants identified complex metabolic networks among IAA, PAA, phenylpropanoid and tryptophan metabolism.
Keywords: Arabidopsis thaliana; aldoxime; auxin; indole-3-acetic acid (IAA); phenylacetic acid (PAA); phenylpropanoid.
© 2021 The Authors New Phytologist © 2021 New Phytologist Foundation.
Figures
References
-
- Andersen MD, Busk PK, Svendsen I, and Møller BL (2000). Cytochromes P-450 from Cassava (Manihot esculentaCrantz) Catalyzing the First Steps in the Biosynthesis of the Cyanogenic Glucosides Linamarin and Lotaustralin: CLONING, FUNCTIONAL EXPRESSION IN PICHIA PASTORIS, AND SUBSTRATE SPECIFICITY OF THE ISOLATED RECOMBINANT ENZYMES*. Journal of Biological Chemistry 275:1966–1975. - PubMed
-
- Aoi Y, Oikawa A, Sasaki R, Huang J, Hayashi K, and Kasahara H (2020b). Arogenate dehydratases can modulate the levels of phenylacetic acid in Arabidopsis. Biochemical and Biophysical Research Communications 524:83–88. - PubMed
-
- Blažević I, Montaut S, Burčul F, Olsen CE, Burow M, Rollin P, and Agerbirk N (2020). Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 169:112100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
