

Studies of aging nonhuman primates illuminate the etiology of early-stage Alzheimer's-like neuropathology: An evolutionary perspective
- PMID: 33960505
- PMCID: PMC8550995
- DOI: 10.1002/ajp.23254
Studies of aging nonhuman primates illuminate the etiology of early-stage Alzheimer's-like neuropathology: An evolutionary perspective
Abstract
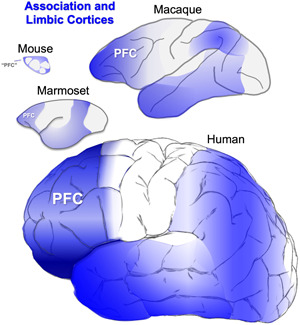
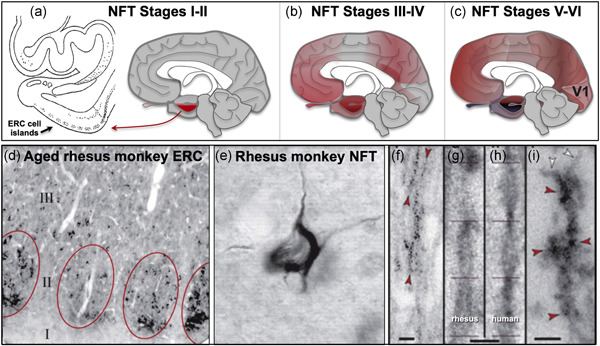
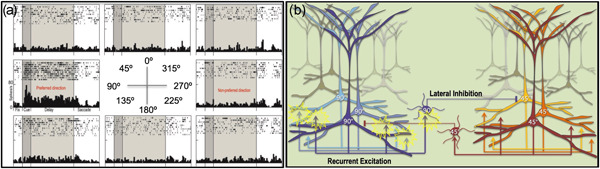
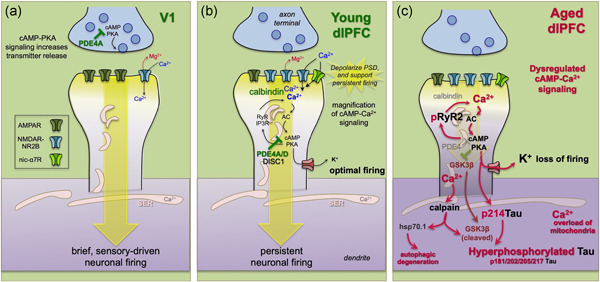
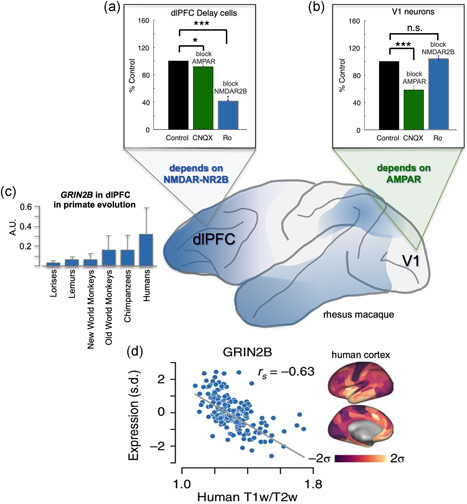
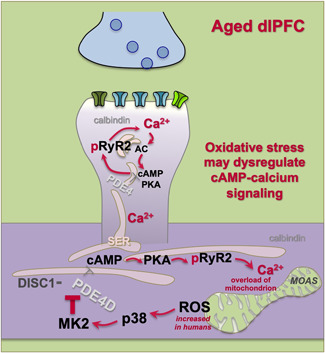
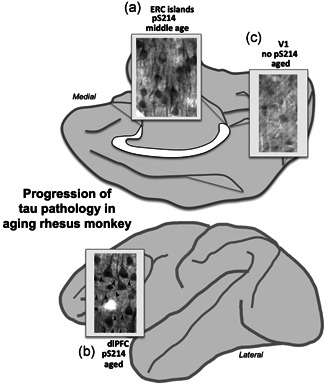
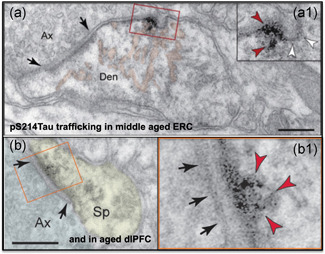
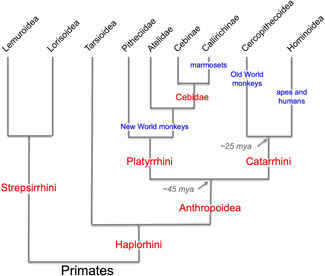
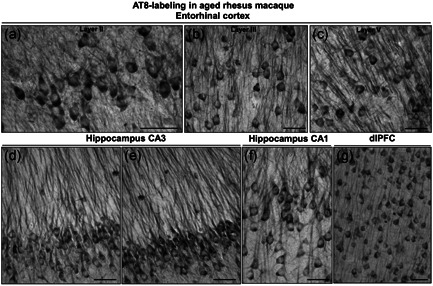
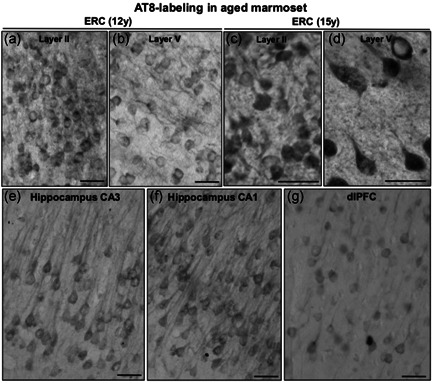
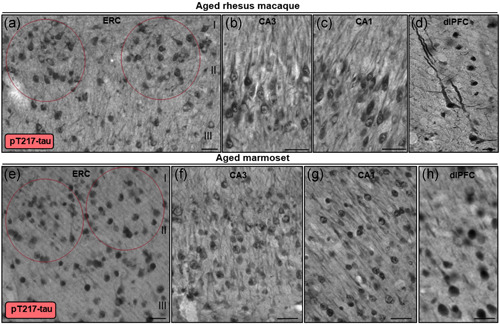
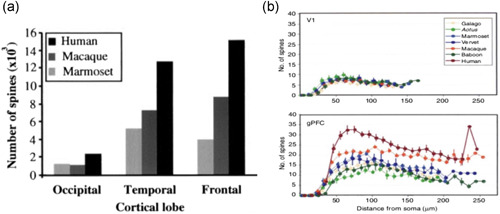
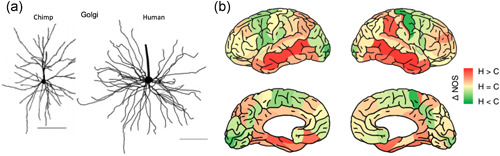
Tau pathology in Alzheimer's disease (AD) preferentially afflicts the limbic and recently enlarged association cortices, causing a progression of mnemonic and cognitive deficits. Although genetic mouse models have helped reveal mechanisms underlying the rare, autosomal-dominant forms of AD, the etiology of the more common, sporadic form of AD remains unknown, and is challenging to study in mice due to their limited association cortex and lifespan. It is also difficult to study in human brains, as early-stage tau phosphorylation can degrade postmortem. In contrast, rhesus monkeys have extensive association cortices, are long-lived, and can undergo perfusion fixation to capture early-stage tau phosphorylation in situ. Most importantly, rhesus monkeys naturally develop amyloid plaques, neurofibrillary tangles comprised of hyperphosphorylated tau, synaptic loss, and cognitive deficits with advancing age, and thus can be used to identify the early molecular events that initiate and propel neuropathology in the aging association cortices. Studies to date suggest that the particular molecular signaling events needed for higher cognition-for example, high levels of calcium to maintain persistent neuronal firing- lead to tau phosphorylation and inflammation when dysregulated with advancing age. The expression of NMDAR-NR2B (GluN2B)-the subunit that fluxes high levels of calcium-increases over the cortical hierarchy and with the expansion of association cortex in primate evolution, consistent with patterns of tau pathology. In the rhesus monkey dorsolateral prefrontal cortex, spines contain NMDAR-NR2B and the molecular machinery to magnify internal calcium release near the synapse, as well as phosphodiesterases, mGluR3, and calbindin to regulate calcium signaling. Loss of regulation with inflammation and/or aging appears to be a key factor in initiating tau pathology. The vast expansion in the numbers of these synapses over primate evolution is consistent with the degree of tau pathology seen across species: marmoset < rhesus monkey < chimpanzee < human, culminating in the vast neurodegeneration seen in humans with AD.
Keywords: AT8; Alzheimer's disease; calcium; entorhinal cortex; p217Tau; prefrontal cortex.
© 2021 The Authors. American Journal of Primatology Published by Wiley Periodicals LLC.
Figures
References
-
- Ardesch, D. J. , Scholtens, L. H. , Li, L. , Preuss, T. M. , Rilling, J. K. , & van den Heuvel, M. P. (2019). Evolutionary expansion of connectivity between multimodal association areas in the human brain compared with chimpanzees. Proceedings of the National Academy of Sciences of the United States of America, 116, 7101–7106. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
