

CD1a selectively captures endogenous cellular lipids that broadly block T cell response
- PMID: 33961028
- PMCID: PMC8111460
- DOI: 10.1084/jem.20202699
CD1a selectively captures endogenous cellular lipids that broadly block T cell response
Abstract
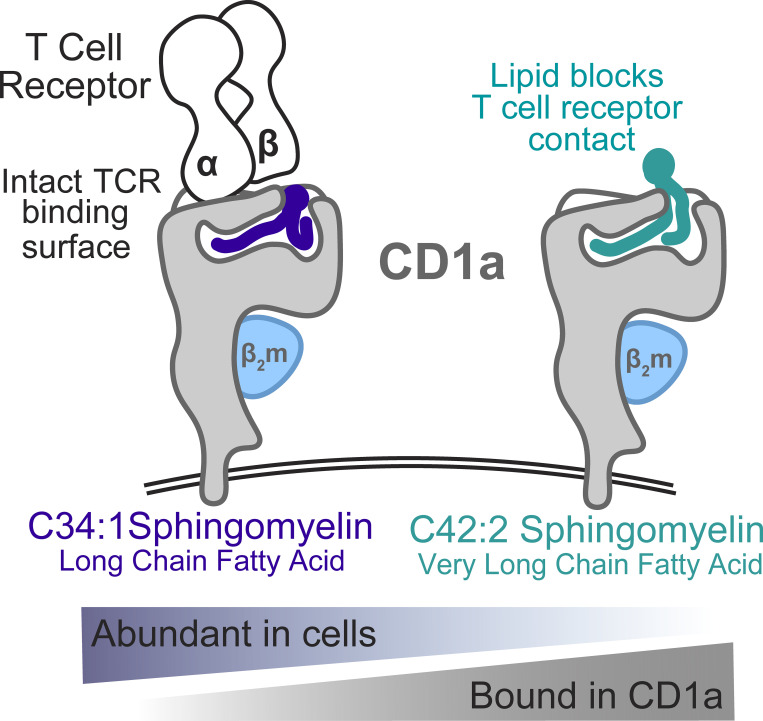
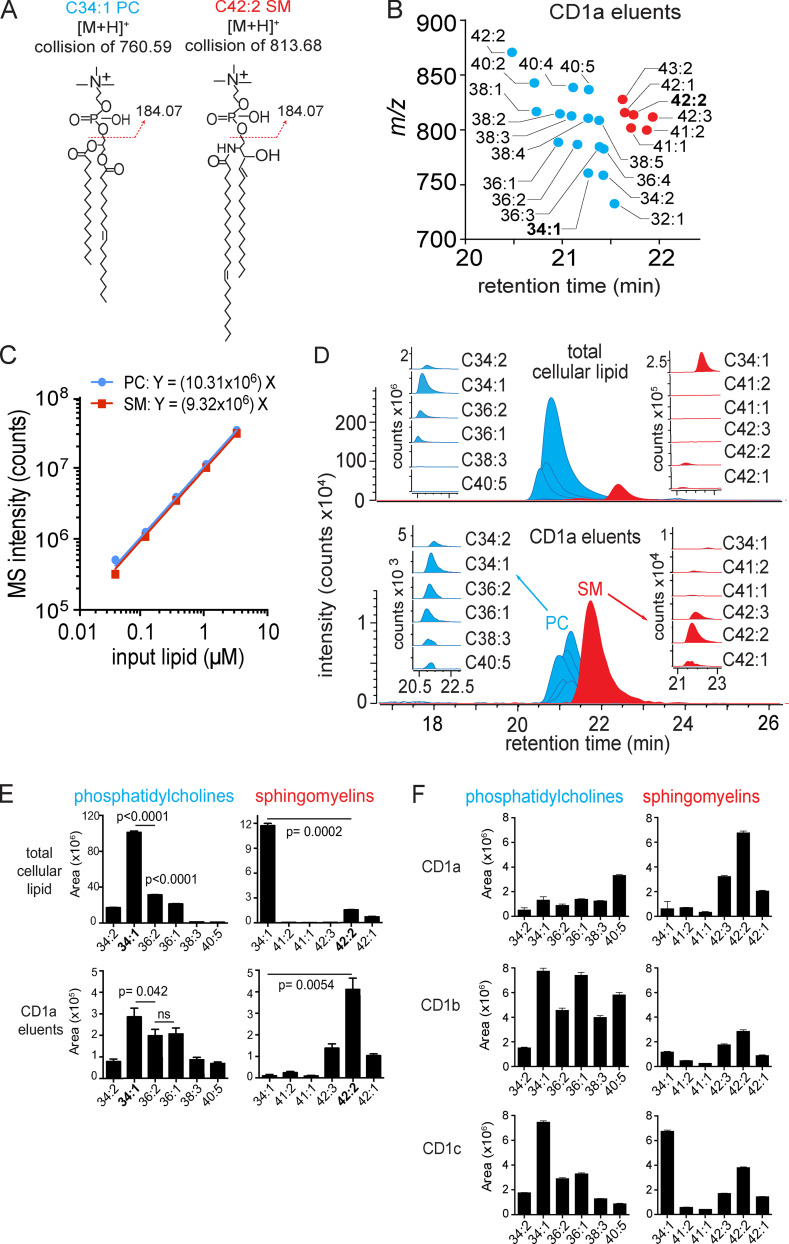
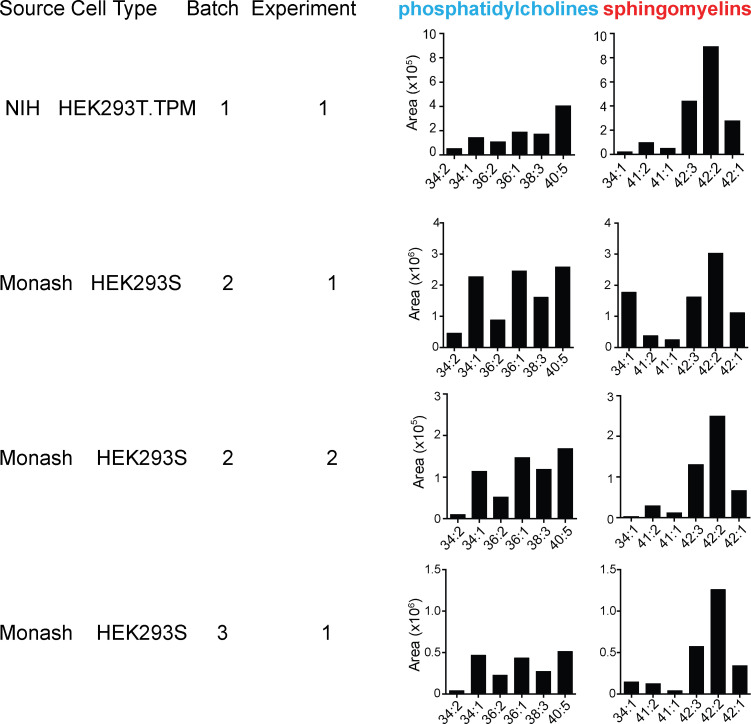
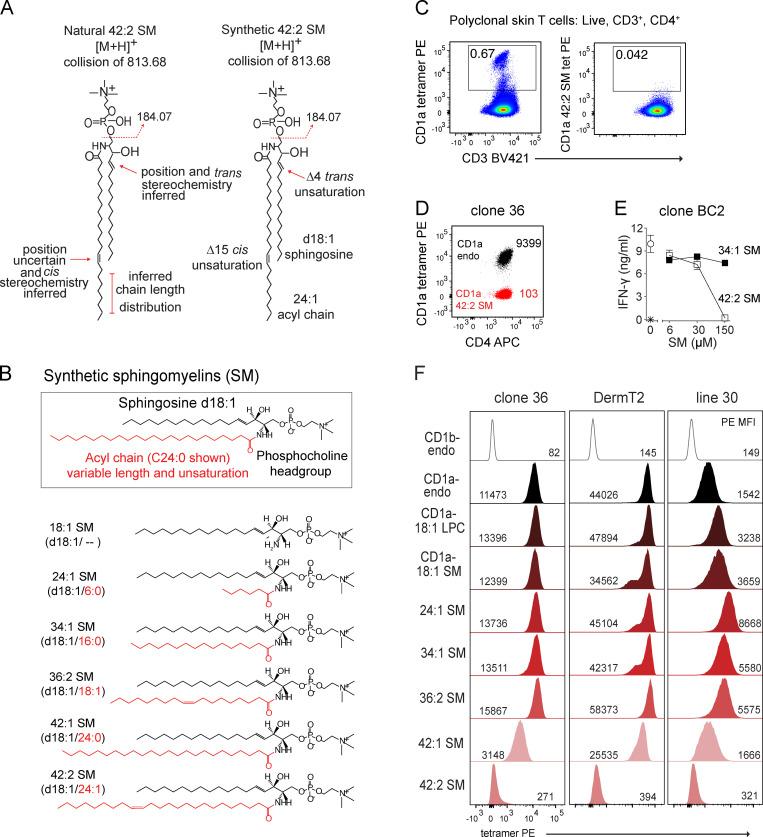
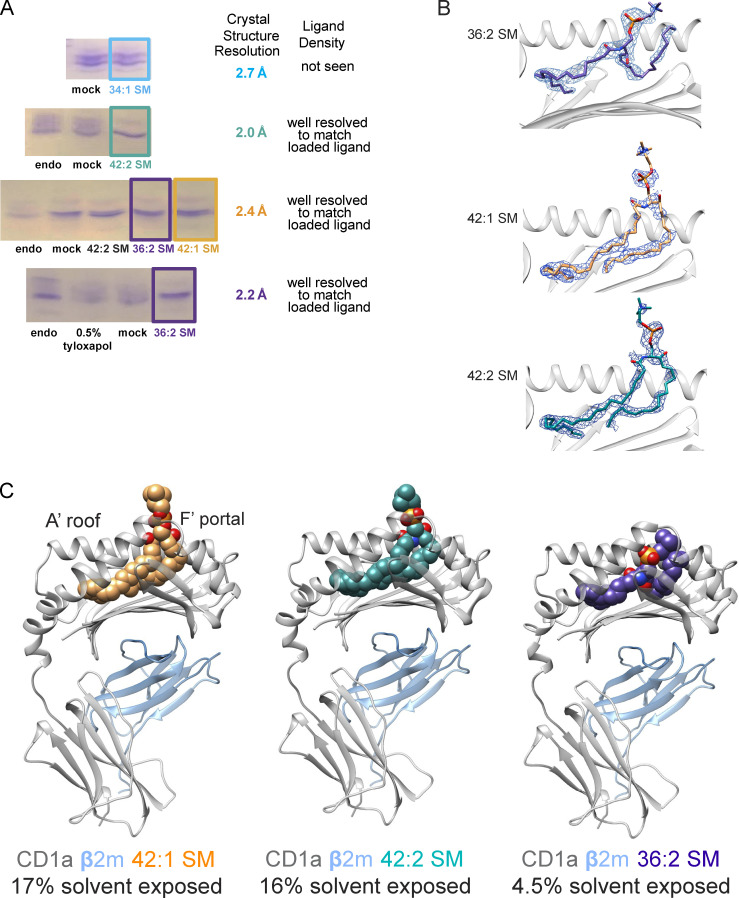
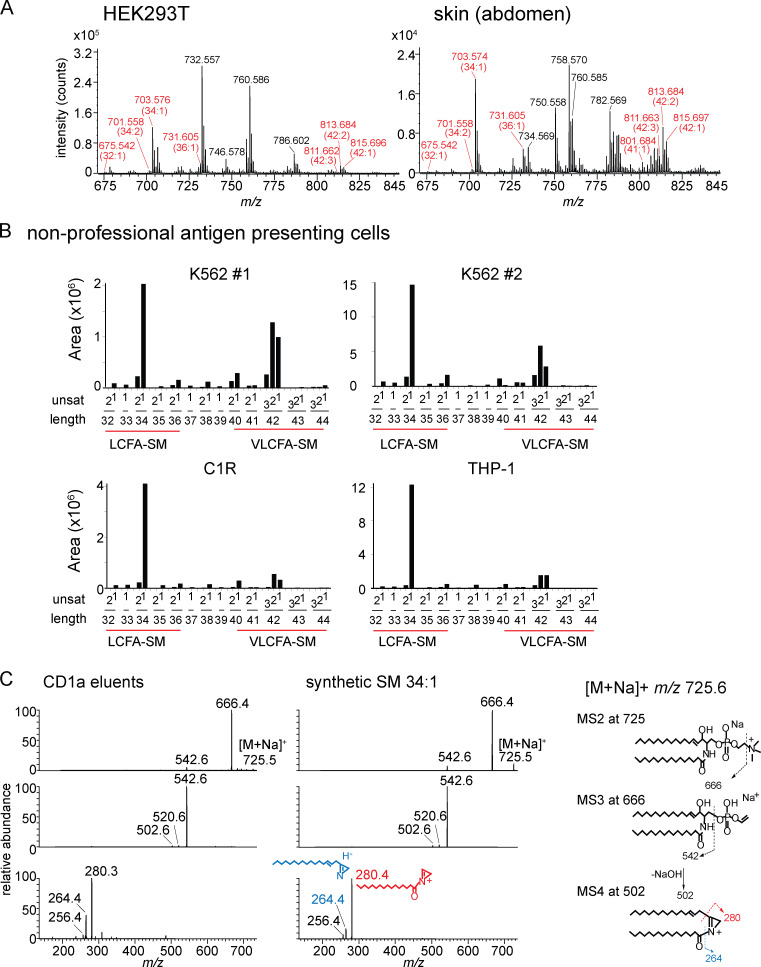
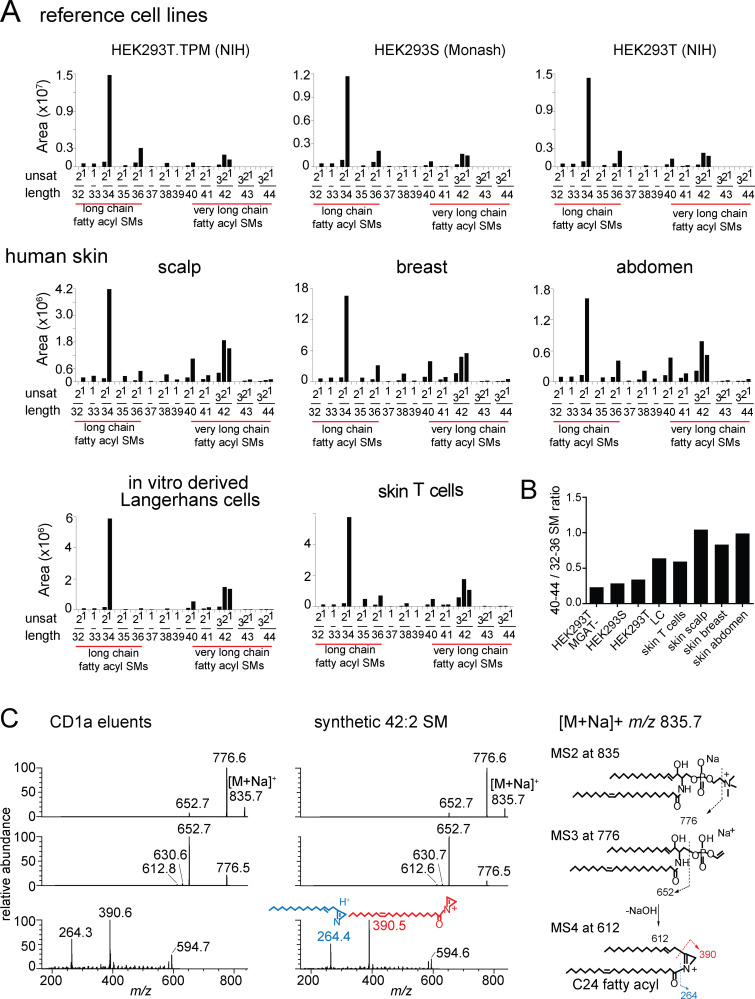
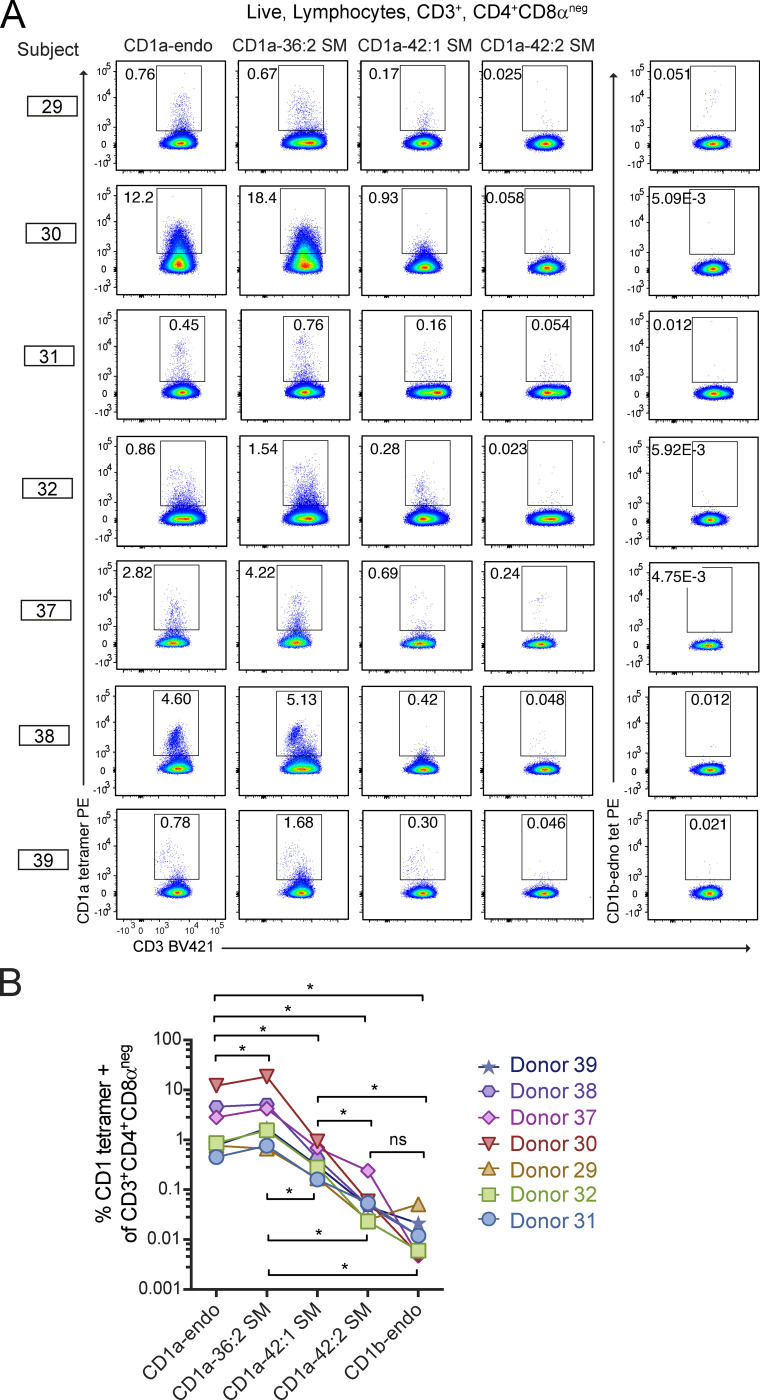
We optimized lipidomics methods to broadly detect endogenous lipids bound to cellular CD1a proteins. Whereas membrane phospholipids dominate in cells, CD1a preferentially captured sphingolipids, especially a C42, doubly unsaturated sphingomyelin (42:2 SM). The natural 42:2 SM but not the more common 34:1 SM blocked CD1a tetramer binding to T cells in all human subjects tested. Thus, cellular CD1a selectively captures a particular endogenous lipid that broadly blocks its binding to TCRs. Crystal structures show that the short cellular SMs stabilized a triad of surface residues to remain flush with CD1a, but the longer lipids forced the phosphocholine group to ride above the display platform to hinder TCR approach. Whereas nearly all models emphasize antigen-mediated T cell activation, we propose that the CD1a system has intrinsic autoreactivity and is negatively regulated by natural endogenous inhibitors selectively bound in its cleft. Further, the detailed chemical structures of natural blockers could guide future design of therapeutic blockers of CD1a response.
© 2021 Cotton et al.
Conflict of interest statement
Disclosures: R.N. Cotton reported personal fees from MPM Capital outside the submitted work. G. Ogg reported grants from UCB outside the submitted work. No other disclosures were reported.
Figures

Comment in
-
CD1a autoreactivity: When size does matter.J Exp Med. 2021 Jul 5;218(7):e20210531. doi: 10.1084/jem.20210531. Epub 2021 May 20. J Exp Med. 2021. PMID: 34014254 Free PMC article.
References
-
- Afonine, P.V., Grosse-Kunstleve R.W., Echols N., Headd J.J., Moriarty N.W., Mustyakimov M., Terwilliger T.C., Urzhumtsev A., Zwart P.H., and Adams P.D.. 2012. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 68:352–367. 10.1107/S0907444912001308 - DOI - PMC - PubMed
-
- Birkinshaw, R.W., Pellicci D.G., Cheng T.Y., Keller A.N., Sandoval-Romero M., Gras S., de Jong A., Uldrich A.P., Moody D.B., Godfrey D.I., and Rossjohn J.. 2015. αβ T cell antigen receptor recognition of CD1a presenting self lipid ligands. Nat. Immunol. 16:258–266. 10.1038/ni.3098 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
