

Iron status influences non-alcoholic fatty liver disease in obesity through the gut microbiome
- PMID: 33962692
- PMCID: PMC8106161
- DOI: 10.1186/s40168-021-01052-7
Iron status influences non-alcoholic fatty liver disease in obesity through the gut microbiome
Abstract
Background: The gut microbiome and iron status are known to play a role in the pathophysiology of non-alcoholic fatty liver disease (NAFLD), although their complex interaction remains unclear.
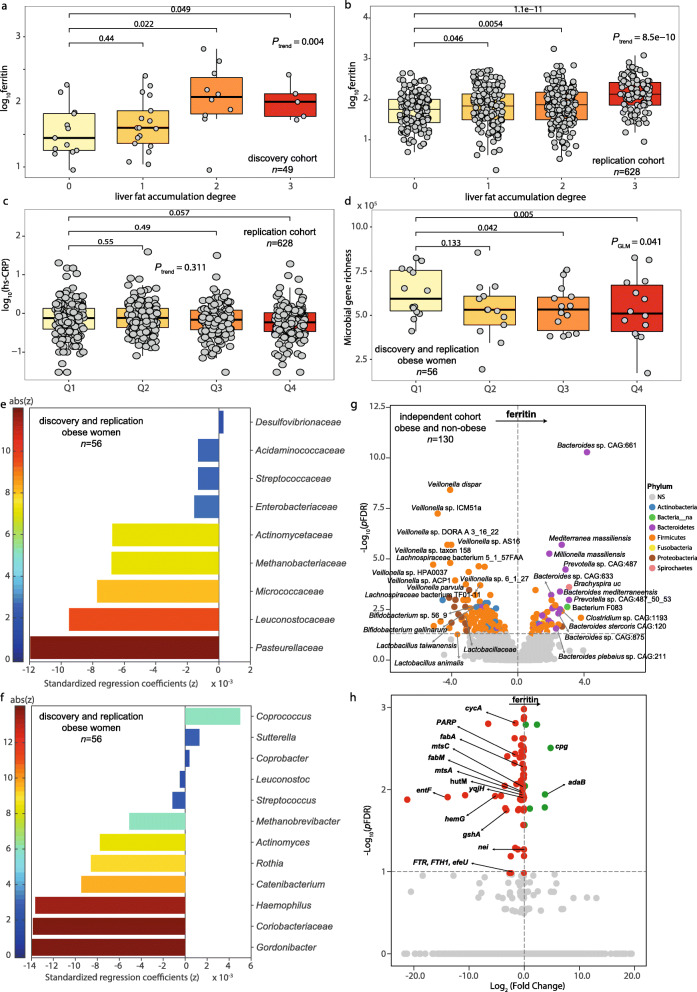
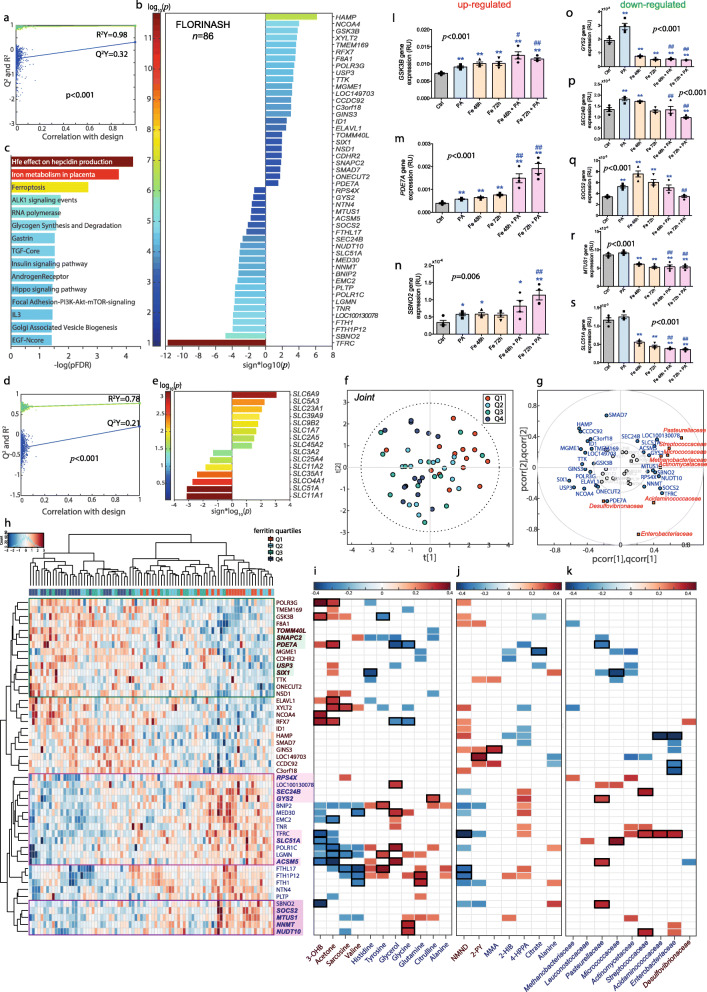
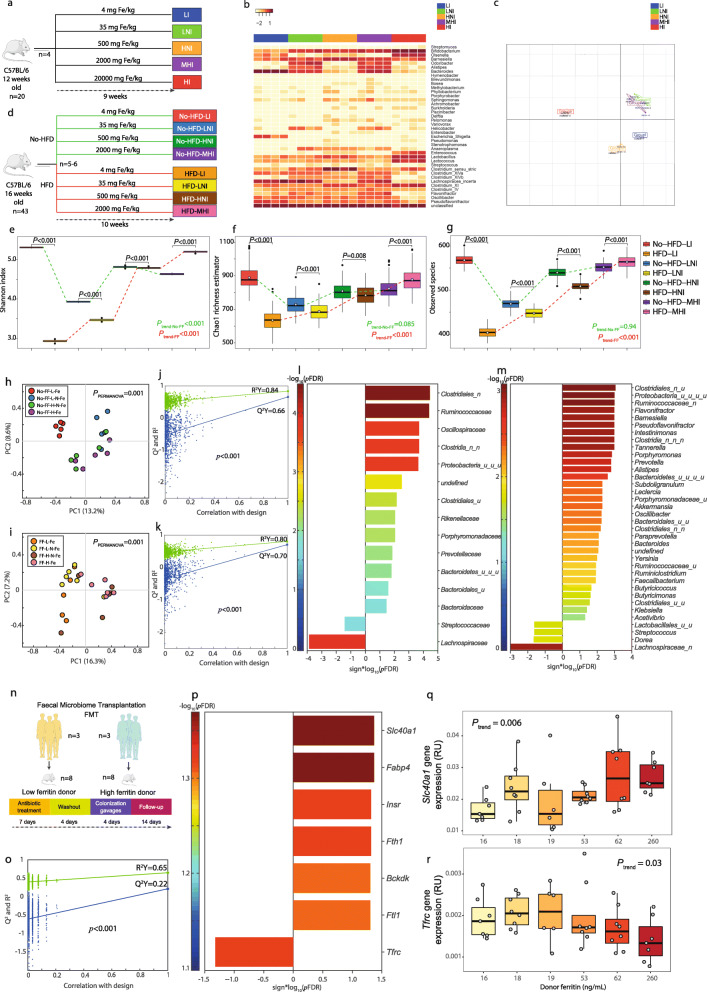
Results: Here, we applied an integrative systems medicine approach (faecal metagenomics, plasma and urine metabolomics, hepatic transcriptomics) in 2 well-characterised human cohorts of subjects with obesity (discovery n = 49 and validation n = 628) and an independent cohort formed by both individuals with and without obesity (n = 130), combined with in vitro and animal models. Serum ferritin levels, as a markers of liver iron stores, were positively associated with liver fat accumulation in parallel with lower gut microbial gene richness, composition and functionality. Specifically, ferritin had strong negative associations with the Pasteurellaceae, Leuconostocaceae and Micrococcaea families. It also had consistent negative associations with several Veillonella, Bifidobacterium and Lactobacillus species, but positive associations with Bacteroides and Prevotella spp. Notably, the ferritin-associated bacterial families had a strong correlation with iron-related liver genes. In addition, several bacterial functions related to iron metabolism (transport, chelation, heme and siderophore biosynthesis) and NAFLD (fatty acid and glutathione biosynthesis) were also associated with the host serum ferritin levels. This iron-related microbiome signature was linked to a transcriptomic and metabolomic signature associated to the degree of liver fat accumulation through hepatic glucose metabolism. In particular, we found a consistent association among serum ferritin, Pasteurellaceae and Micrococcacea families, bacterial functions involved in histidine transport, the host circulating histidine levels and the liver expression of GYS2 and SEC24B. Serum ferritin was also related to bacterial glycine transporters, the host glycine serum levels and the liver expression of glycine transporters. The transcriptomic findings were replicated in human primary hepatocytes, where iron supplementation also led to triglycerides accumulation and induced the expression of lipid and iron metabolism genes in synergy with palmitic acid. We further explored the direct impact of the microbiome on iron metabolism and liver fact accumulation through transplantation of faecal microbiota into recipient's mice. In line with the results in humans, transplantation from 'high ferritin donors' resulted in alterations in several genes related to iron metabolism and fatty acid accumulation in recipient's mice.
Conclusions: Altogether, a significant interplay among the gut microbiome, iron status and liver fat accumulation is revealed, with potential significance for target therapies. Video abstract.
Keywords: Ferritin; Gut microbiome; Iron status; Metagenomics; Non-alcoholic fatty liver disease; Obesity; Shotgun sequencing; Systems medicine.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
