

ZmFdC2 Encoding a Ferredoxin Protein With C-Terminus Extension Is Indispensable for Maize Growth
- PMID: 33968104
- PMCID: PMC8104031
- DOI: 10.3389/fpls.2021.646359
ZmFdC2 Encoding a Ferredoxin Protein With C-Terminus Extension Is Indispensable for Maize Growth
Abstract
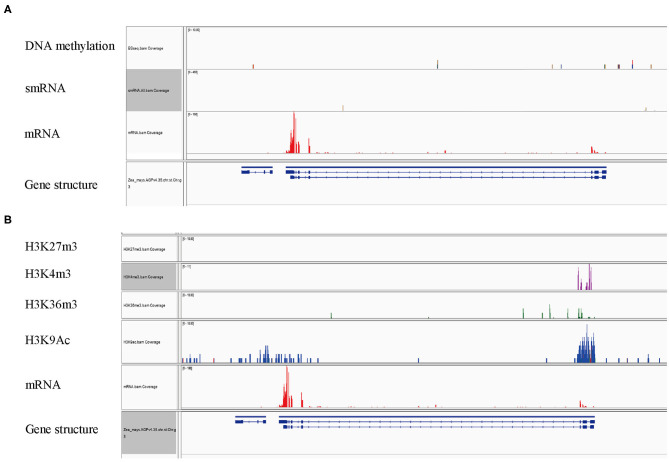
As important electron carriers, ferredoxin (Fd) proteins play important roles in photosynthesis, and the assimilation of CO2, nitrate, sulfate, and other metabolites. In addition to the well-studied Fds, plant genome encodes two Fd-like protein members named FdC1 and FdC2, which have extension regions at the C-terminus of the 2Fe-2S cluster. Mutation or overexpression of FdC genes caused alterations in photosynthetic electron transfer rate in rice and Arabidopsis. Maize genome contains one copy of each FdC gene. However, the functions of these genes have not been reported. In this study, we identified the ZmFdC2 gene by forward genetics approach. Mutation of this gene causes impaired photosynthetic electron transport and collapsed chloroplasts. The mutant plant is seedling-lethal, indicating the indispensable function of ZmFdC2 gene in maize development. The ZmFdC2 gene is specifically expressed in photosynthetic tissues and induced by light treatment, and the encoded protein is localized on chloroplast, implying its specialized function in photosynthesis. Furthermore, ZmFdC2 expression was detected in both mesophyll cells and bundle sheath cells, the two cell types specialized for C4 and C3 photosynthesis pathways in maize. Epigenomic analyses showed that ZmFdC2 locus was enriched for active histone modifications. Our results demonstrate that ZmFdC2 is a key component of the photosynthesis pathway and is crucial for the development of maize.
Keywords: ZmFdC2; electron transfer; growth; maize; photosynthesis.
Copyright © 2021 Chen, Zhong, Yang, Zhao, Dai, Zeng, Wang, Gao and Li.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The reviewer YW declared a shared affiliation, with no collaboration, with several of the authors, SL and YZ, to the handling editor at the time of the review.
Figures






Similar articles
-
Mutation of FdC2 gene encoding a ferredoxin-like protein with C-terminal extension causes yellow-green leaf phenotype in rice.Plant Sci. 2015 Sep;238:127-34. doi: 10.1016/j.plantsci.2015.06.010. Epub 2015 Jun 17. Plant Sci. 2015. PMID: 26259181
-
Functional Inactivation of Putative Photosynthetic Electron Acceptor Ferredoxin C2 (FdC2) Induces Delayed Heading Date and Decreased Photosynthetic Rate in Rice.PLoS One. 2015 Nov 24;10(11):e0143361. doi: 10.1371/journal.pone.0143361. eCollection 2015. PLoS One. 2015. PMID: 26598971 Free PMC article.
-
The promoter of rbcS in a C3 plant (rice) directs organ-specific, light-dependent expression in a C4 plant (maize), but does not confer bundle sheath cell-specific expression.Plant Mol Biol. 2000 Sep;44(1):99-106. doi: 10.1023/a:1006461812053. Plant Mol Biol. 2000. PMID: 11094984
-
Plastid transport and metabolism of C3 and C4 plants--comparative analysis and possible biotechnological exploitation.Curr Opin Plant Biol. 2010 Jun;13(3):257-65. doi: 10.1016/j.pbi.2010.01.007. Epub 2010 Feb 24. Curr Opin Plant Biol. 2010. PMID: 20188622 Review.
-
Cell-type-specific differentiation of chloroplasts in C4 plants.Trends Plant Sci. 2009 Feb;14(2):100-9. doi: 10.1016/j.tplants.2008.11.006. Epub 2009 Jan 21. Trends Plant Sci. 2009. PMID: 19162526 Review.
Cited by
-
LAZARUS 1 functions as a positive regulator of plant immunity and systemic acquired resistance.Front Plant Sci. 2024 Nov 20;15:1490466. doi: 10.3389/fpls.2024.1490466. eCollection 2024. Front Plant Sci. 2024. PMID: 39634069 Free PMC article.
-
Ferredoxin C2 is required for chlorophyll biosynthesis and accumulation of photosynthetic antennae in Arabidopsis.Plant Cell Environ. 2023 Nov;46(11):3287-3304. doi: 10.1111/pce.14667. Epub 2023 Jul 10. Plant Cell Environ. 2023. PMID: 37427830 Free PMC article.
-
Enhancing Photosynthesis and Plant Productivity through Genetic Modification.Cells. 2024 Aug 7;13(16):1319. doi: 10.3390/cells13161319. Cells. 2024. PMID: 39195209 Free PMC article. Review.
-
Comprehensive identification, stress responses and protein-protein interactions of the ferredoxin gene family in the fast-growing tree species Paulownia fortunei.BMC Plant Biol. 2025 Jul 29;25(1):975. doi: 10.1186/s12870-025-06909-9. BMC Plant Biol. 2025. PMID: 40730946 Free PMC article.
References
-
- Blanco N. E., Ceccoli R. D., Via M. V., Voss I., Segretin M. E., Bravo-Almonacid F. F., et al. . (2013). Expression of the minor isoform pea ferredoxin in tobacco alters photosynthetic electron partitioning and enhances cyclic electron flow. Plant Physiol. 161, 866–879. 10.1104/pp.112.211078 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous