

Biallelic variants in HPDL cause pure and complicated hereditary spastic paraplegia
- PMID: 33970200
- PMCID: PMC8219359
- DOI: 10.1093/brain/awab041
Biallelic variants in HPDL cause pure and complicated hereditary spastic paraplegia
Erratum in
-
Erratum to: Biallelic variants in HPDL cause pure and complicated hereditary spastic paraplegia.Brain. 2021 Sep 4;144(8):e70. doi: 10.1093/brain/awab193. Brain. 2021. PMID: 34480796 Free PMC article. No abstract available.
Abstract
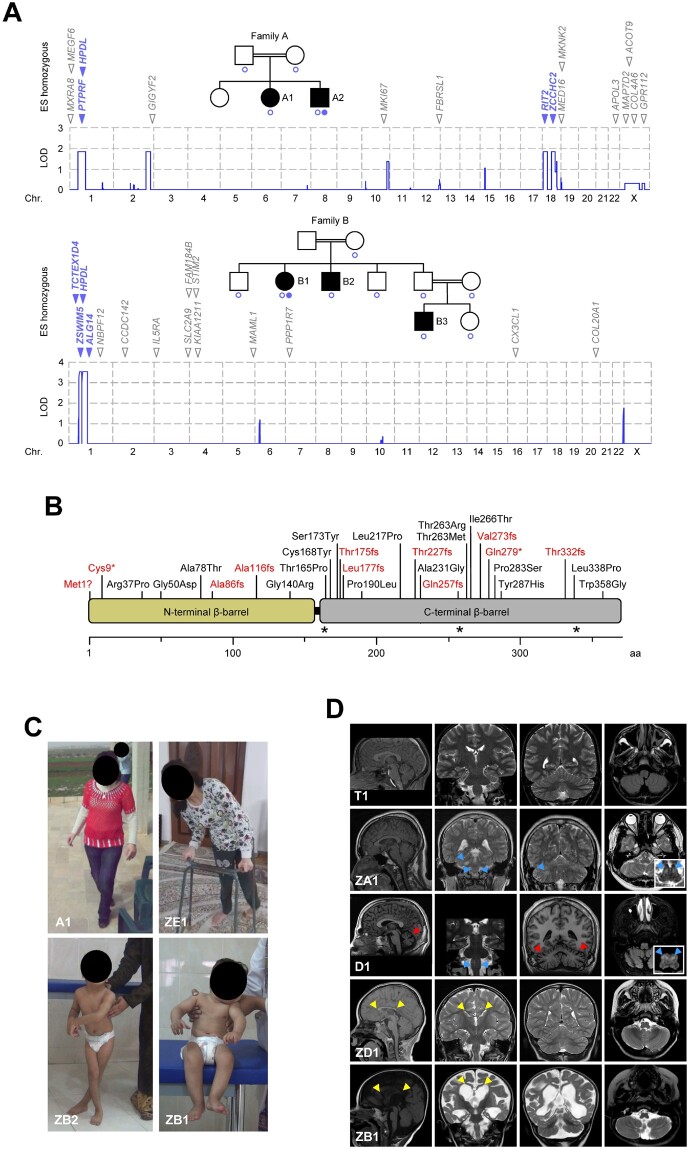
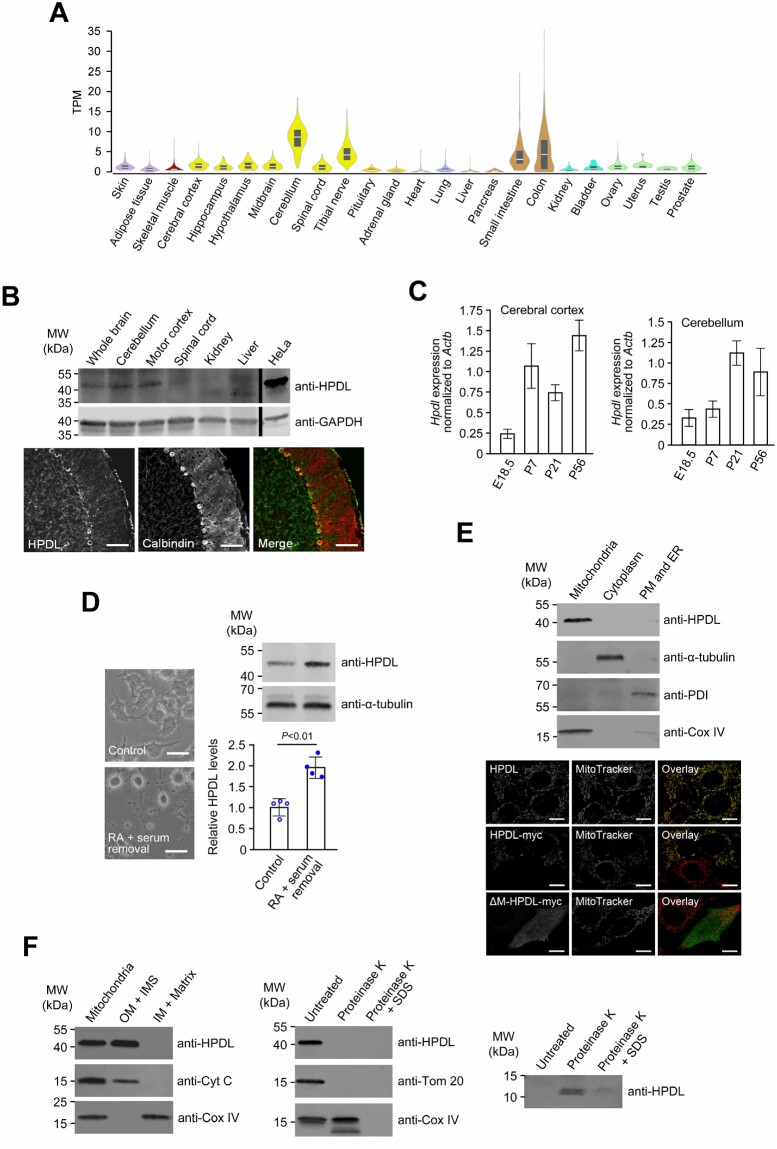
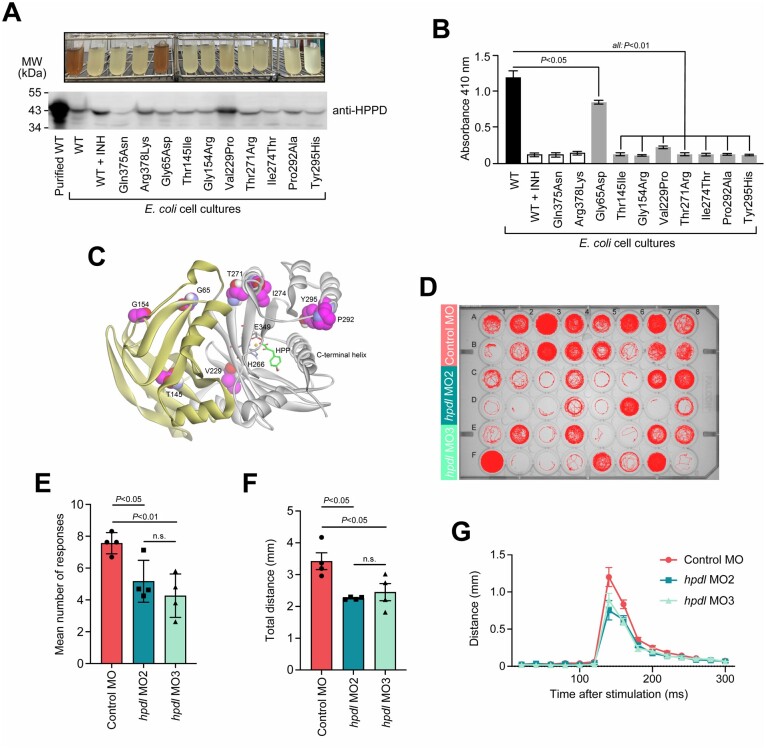
Human 4-hydroxyphenylpyruvate dioxygenase-like (HPDL) is a putative iron-containing non-heme oxygenase of unknown specificity and biological significance. We report 25 families containing 34 individuals with neurological disease associated with biallelic HPDL variants. Phenotypes ranged from juvenile-onset pure hereditary spastic paraplegia to infantile-onset spasticity and global developmental delays, sometimes complicated by episodes of neurological and respiratory decompensation. Variants included bona fide pathogenic truncating changes, although most were missense substitutions. Functionality of variants could not be determined directly as the enzymatic specificity of HPDL is unknown; however, when HPDL missense substitutions were introduced into 4-hydroxyphenylpyruvate dioxygenase (HPPD, an HPDL orthologue), they impaired the ability of HPPD to convert 4-hydroxyphenylpyruvate into homogentisate. Moreover, three additional sets of experiments provided evidence for a role of HPDL in the nervous system and further supported its link to neurological disease: (i) HPDL was expressed in the nervous system and expression increased during neural differentiation; (ii) knockdown of zebrafish hpdl led to abnormal motor behaviour, replicating aspects of the human disease; and (iii) HPDL localized to mitochondria, consistent with mitochondrial disease that is often associated with neurological manifestations. Our findings suggest that biallelic HPDL variants cause a syndrome varying from juvenile-onset pure hereditary spastic paraplegia to infantile-onset spastic tetraplegia associated with global developmental delays.
Keywords: HPDL; HSP; autosomal recessive; hereditary spastic paraplegia; mitochondrial disorder.
© The Author(s) (2021). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Harding AE. Classification of the hereditary ataxias and paraplegias. Lancet. 1983;321:1151–1155. - PubMed
-
- Hedera P. Hereditary spastic paraplegia overview. In: Adam MP, Ardinger HH, Pagon RA, et al., editors. GeneReviews ®. Seattle (WA): University of Washington, Seattle; 2000. - PubMed
-
- Shribman S, Reid E, Crosby AH, Houlden H, Warner TT.. Hereditary spastic paraplegia: From diagnosis to emerging therapeutic approaches. Lancet Neurol. 2019;18:1136–1146. - PubMed
-
- Casari G, De Fusco M, Ciarmatori S, et al.Spastic paraplegia and OXPHOS impairment caused by mutations in paraplegin, a nuclear-encoded mitochondrial metalloprotease. Cell. 1998;93:973–983. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
