

Oxytocin excites BNST interneurons and inhibits BNST output neurons to the central amygdala
- PMID: 33971215
- PMCID: PMC8297366
- DOI: 10.1016/j.neuropharm.2021.108601
Oxytocin excites BNST interneurons and inhibits BNST output neurons to the central amygdala
Abstract
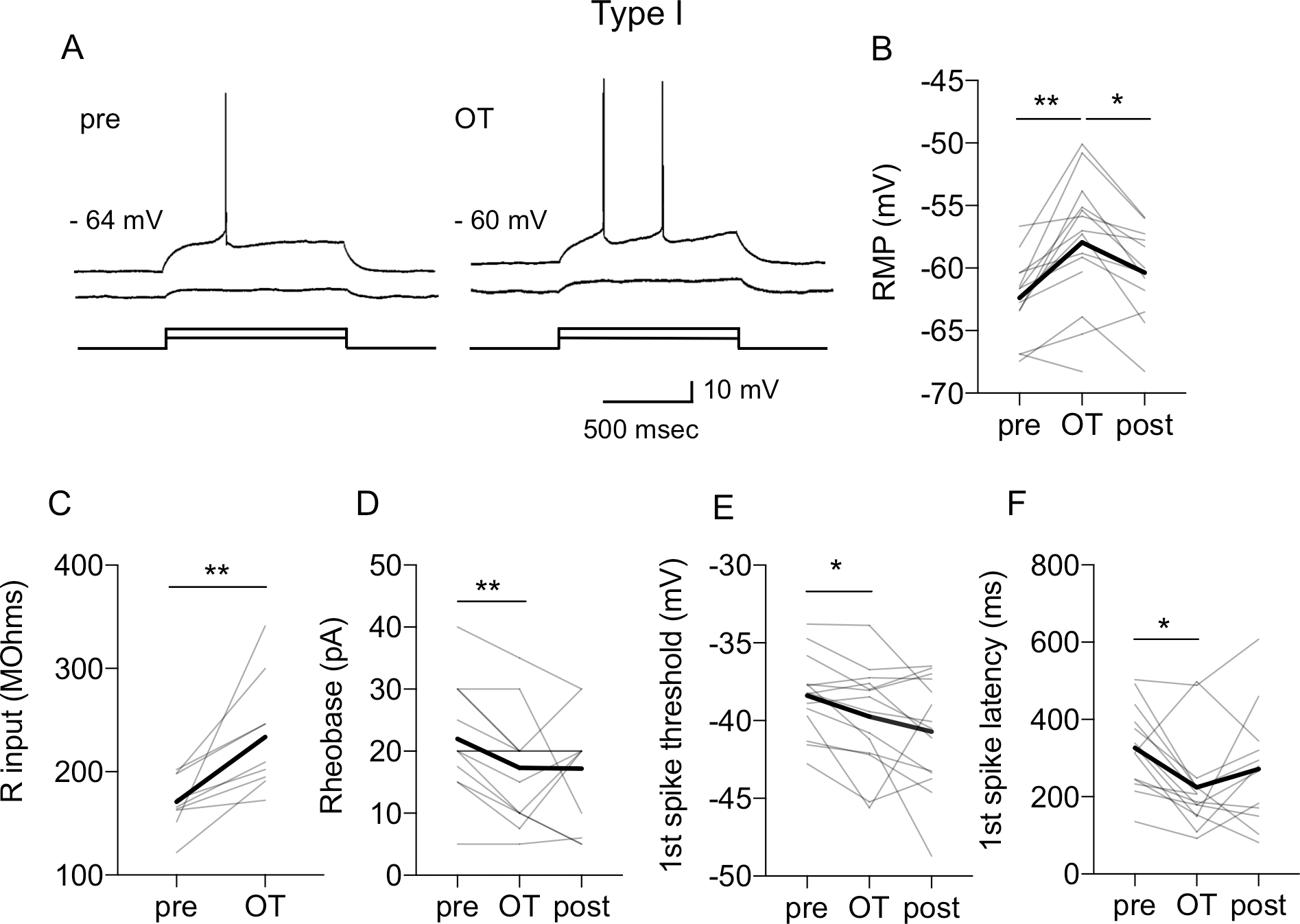
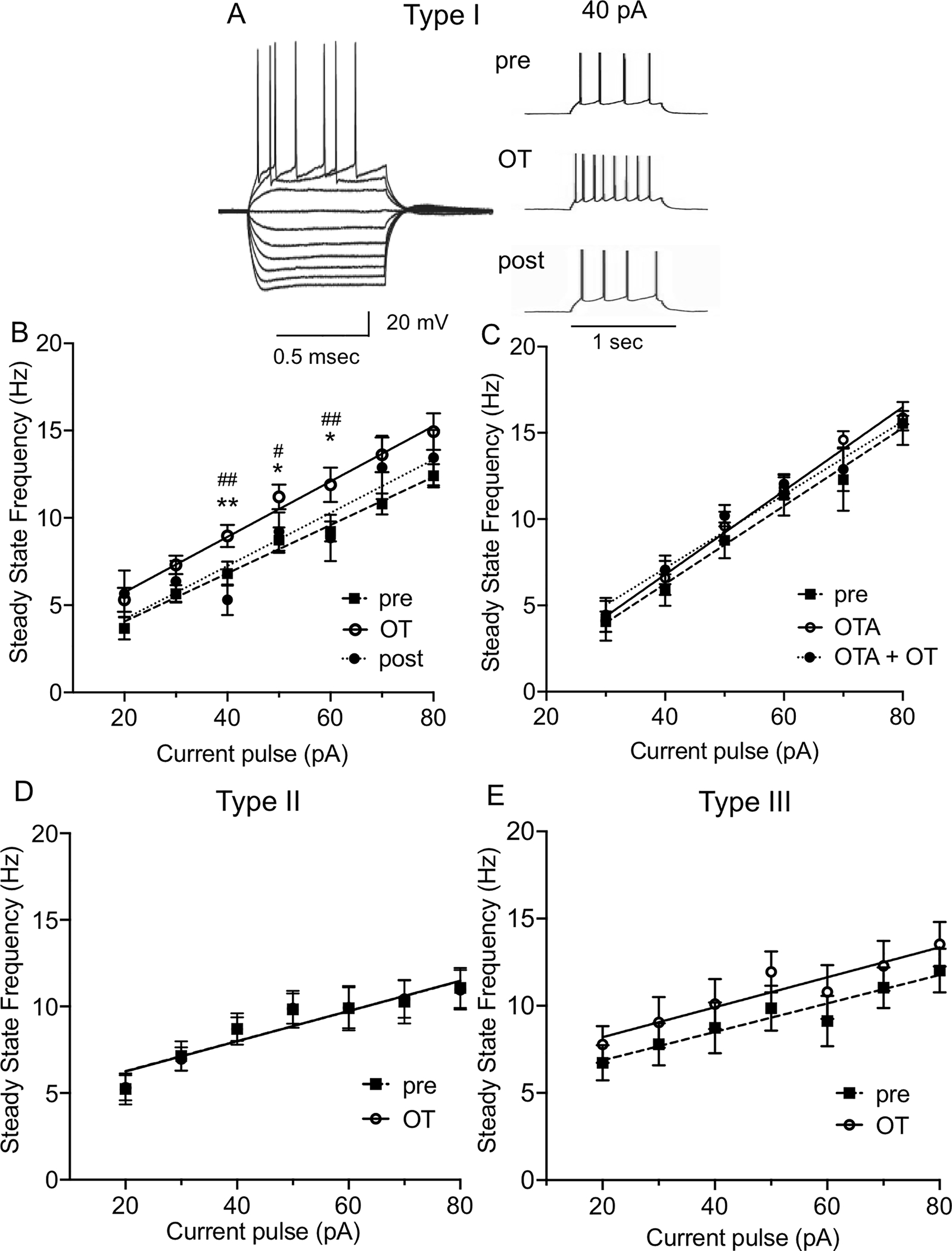
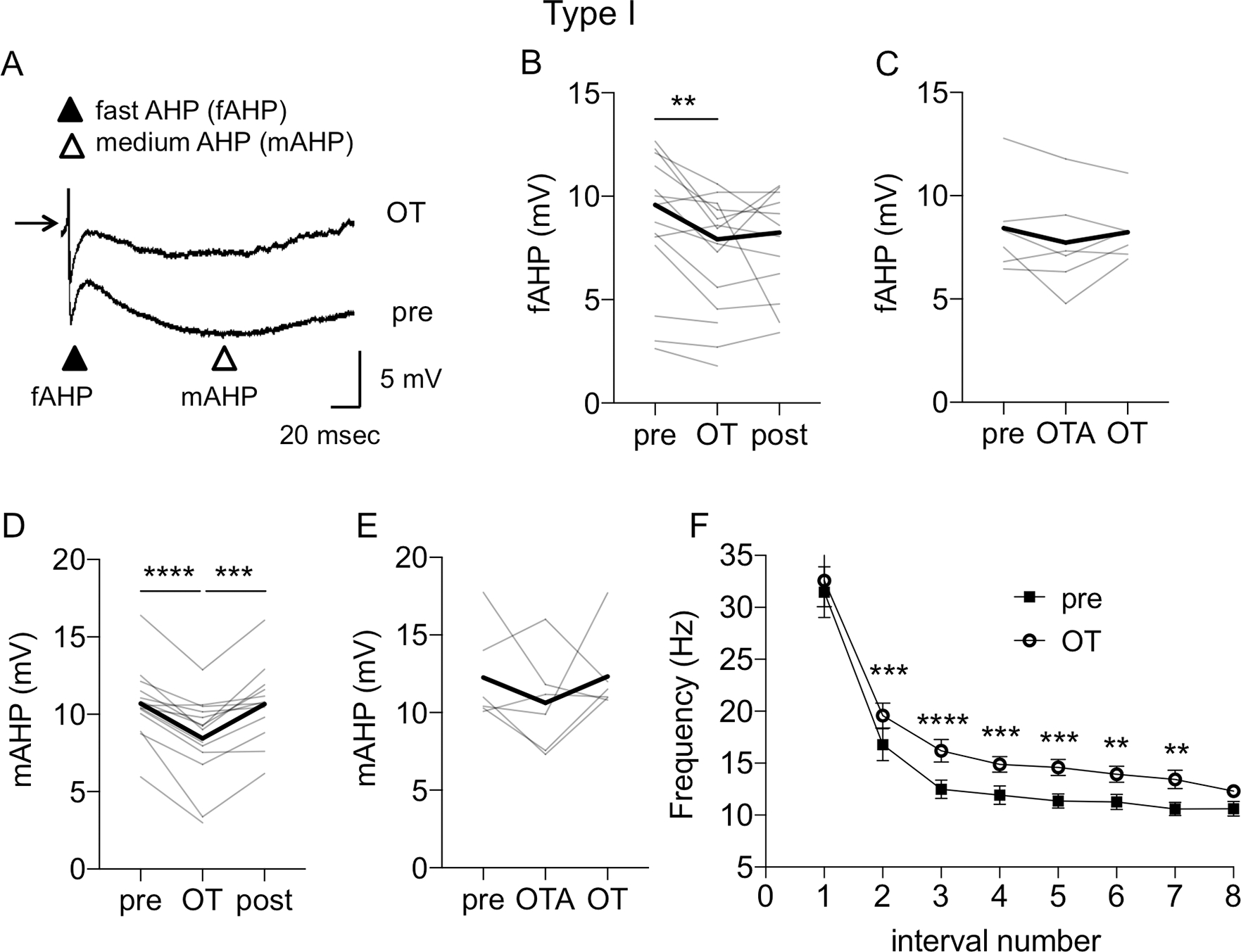
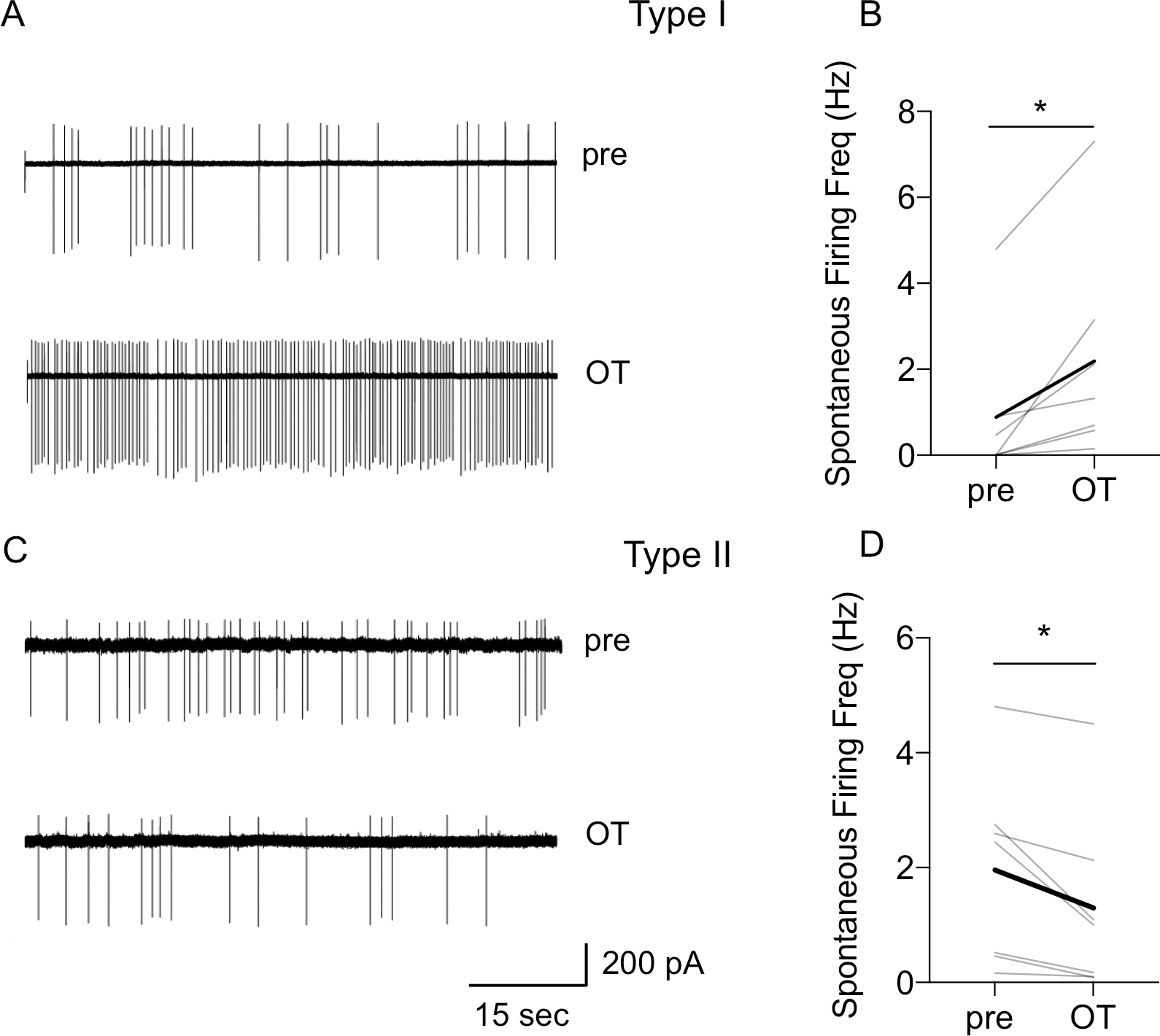
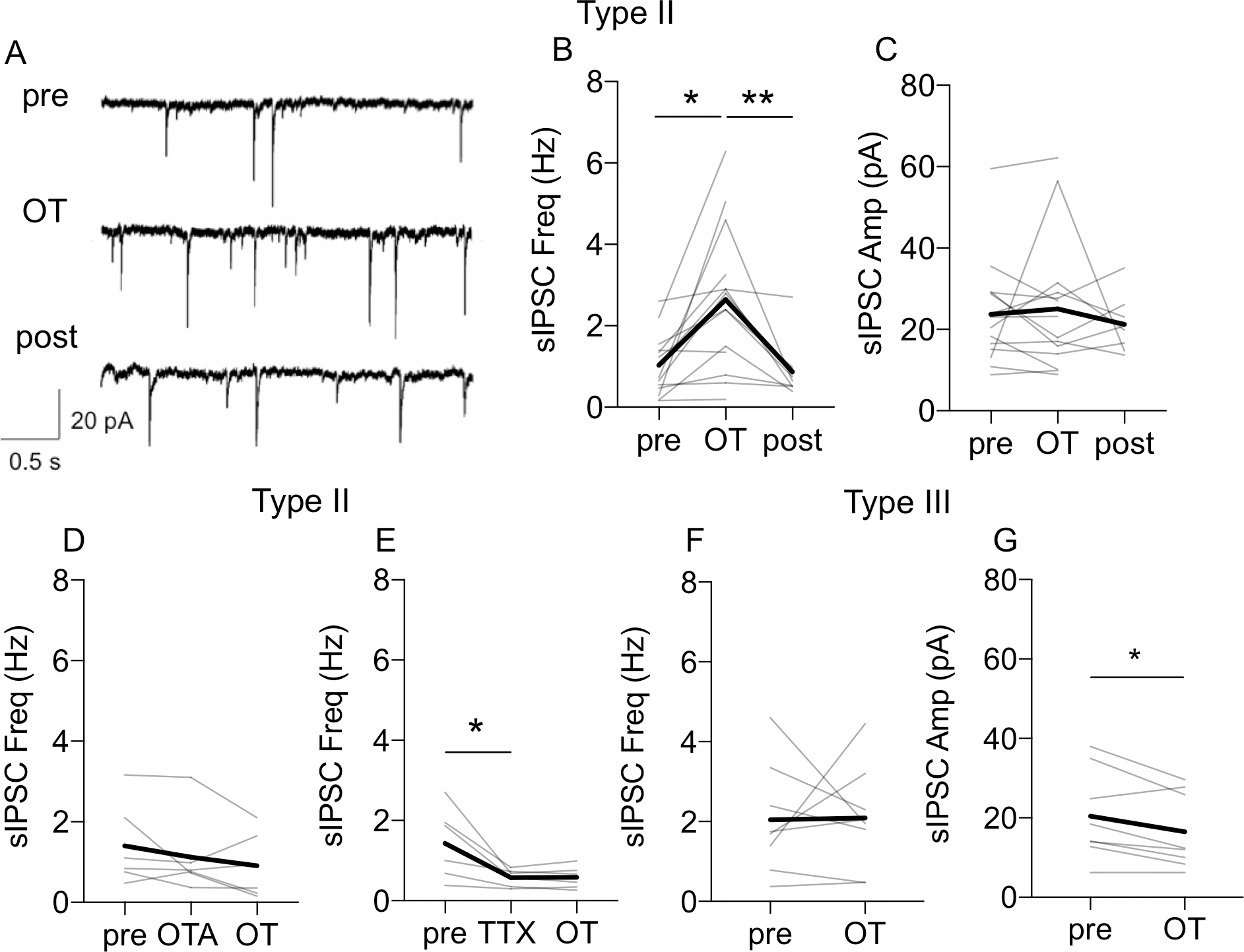
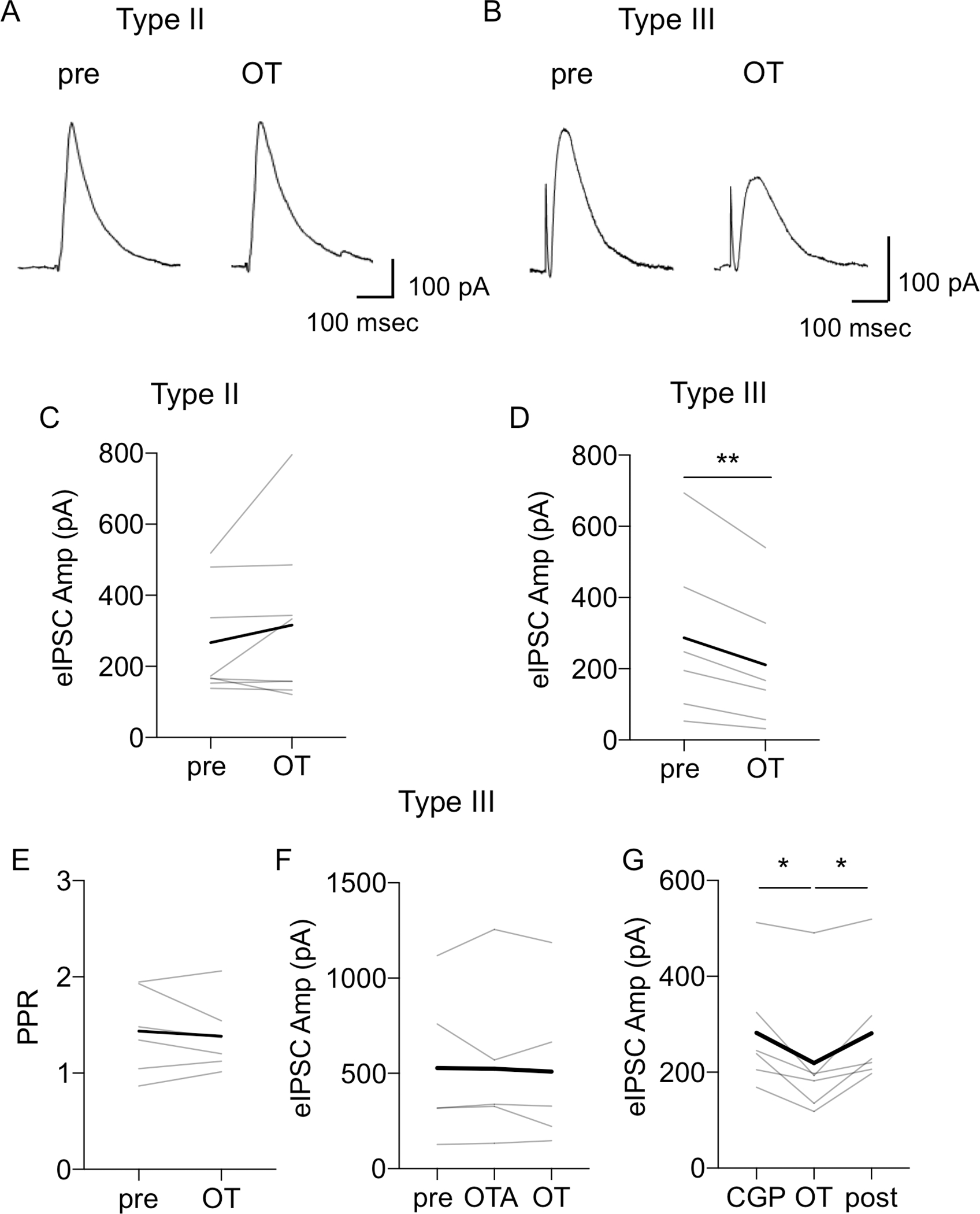
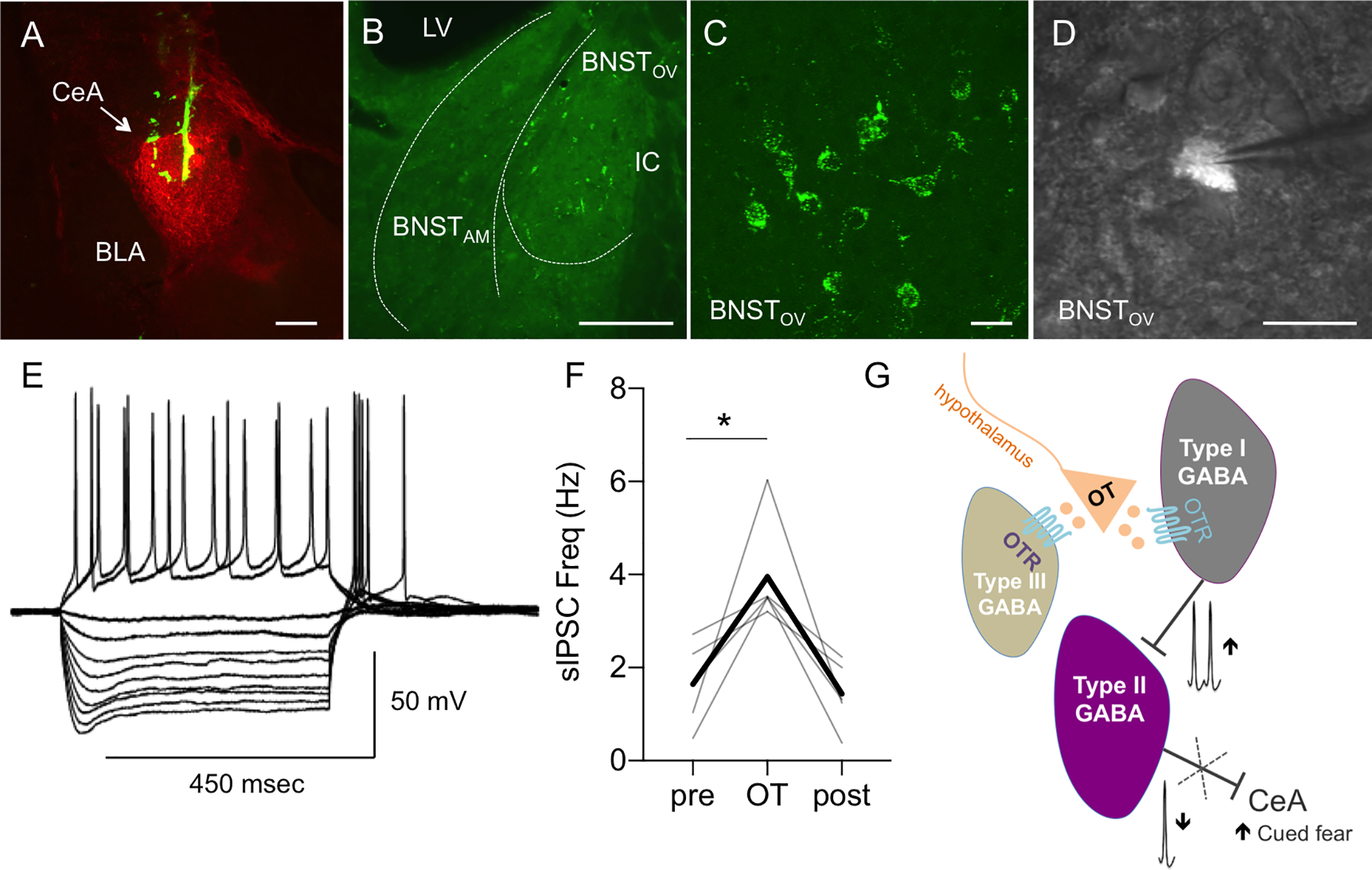
The dorsolateral bed nucleus of the stria terminalis (BNSTDL) has high expression of oxytocin (OT) receptors (OTR), which were shown to facilitate cued fear. However, the role of OTR in the modulation of BNSTDL activity remains elusive. BNSTDL contains GABA-ergic neurons classified based on intrinsic membrane properties into three types. Using in vitro patch-clamp recordings in male rats, we demonstrate that OT selectively excites and increases spontaneous firing rate of Type I BNSTDL neurons. As a consequence, OT increases the frequency, but not amplitude, of spontaneous inhibitory post-synaptic currents (sIPSCs) selectively in Type II neurons, an effect abolished by OTR antagonist or tetrodotoxin, and reduces spontaneous firing rate in these neurons. These results suggest an indirect effect of OT in Type II neurons, which is mediated via OT-induced increase in firing of Type I interneurons. As Type II BNSTDL neurons were shown projecting to the central amygdala (CeA), we also recorded from retrogradely labeled BNST→CeA neurons and we show that OT increases the frequency of sIPSC in these Type II BNST→CeA output neurons. In contrast, in Type III neurons, OT reduces the amplitude, but not frequency, of both sIPSCs and evoked IPSCs via a postsynaptic mechanism without changing their intrinsic excitability. We present a model of fine-tuned modulation of BNSTDL activity by OT, which selectively excites BNSTDL interneurons and inhibits Type II BNST→CeA output neurons. These results suggest that OTR in the BNST might facilitate cued fear by inhibiting the BNST→CeA neurons.
Keywords: BNST; Cell-attached; Central amygdala; Electrophysiology; Oxytocin; Patch-clamp.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Financial Disclosures
The authors declare no conflict of interest and have no financial interests to disclose. Dr. Joanna Dabrowska reports submission of a provisional patent application entitled: Method and System for Testing for Stress-related Mental Disorders, Susceptibility, Disease Progression, Disease Modulation and Treatment Efficacy (# 62/673447).
Figures
Similar articles
-
Oxytocin receptors in the dorsolateral bed nucleus of the stria terminalis (BNST) bias fear learning toward temporally predictable cued fear.Transl Psychiatry. 2019 Apr 18;9(1):140. doi: 10.1038/s41398-019-0474-x. Transl Psychiatry. 2019. PMID: 31000694 Free PMC article.
-
Oxytocin receptor neurotransmission in the dorsolateral bed nucleus of the stria terminalis facilitates the acquisition of cued fear in the fear-potentiated startle paradigm in rats.Neuropharmacology. 2017 Jul 15;121:130-139. doi: 10.1016/j.neuropharm.2017.04.039. Epub 2017 Apr 26. Neuropharmacology. 2017. PMID: 28456687 Free PMC article.
-
Optogenetic study of the projections from the bed nucleus of the stria terminalis to the central amygdala.J Neurophysiol. 2015 Nov;114(5):2903-11. doi: 10.1152/jn.00677.2015. Epub 2015 Sep 23. J Neurophysiol. 2015. PMID: 26400259 Free PMC article.
-
Electrophysiological effects of oxytocin within the bed nuclei of the stria terminalis: influence of reproductive stage and ovarian steroids.Prog Brain Res. 1998;119:321-34. doi: 10.1016/s0079-6123(08)61578-2. Prog Brain Res. 1998. PMID: 10074797 Review.
-
Oxytocin facilitates adaptive fear and attenuates anxiety responses in animal models and human studies-potential interaction with the corticotropin-releasing factor (CRF) system in the bed nucleus of the stria terminalis (BNST).Cell Tissue Res. 2019 Jan;375(1):143-172. doi: 10.1007/s00441-018-2889-8. Epub 2018 Jul 28. Cell Tissue Res. 2019. PMID: 30054732 Free PMC article. Review.
Cited by
-
Intrahypothalamic effects of oxytocin on PVN CRH neurons in response to acute stress.Curr Opin Endocr Metab Res. 2022 Oct;26:100382. doi: 10.1016/j.coemr.2022.100382. Epub 2022 Jul 19. Curr Opin Endocr Metab Res. 2022. PMID: 36618014 Free PMC article.
-
Neural Functions of Hypothalamic Oxytocin and its Regulation.ASN Neuro. 2022 Jan-Dec;14:17590914221100706. doi: 10.1177/17590914221100706. ASN Neuro. 2022. PMID: 35593066 Free PMC article. Review.
-
Paraventricular oxytocin neurons impact energy intake and expenditure: projections to the bed nucleus of the stria terminalis reduce sucrose consumption.Front Endocrinol (Lausanne). 2024 Sep 2;15:1449326. doi: 10.3389/fendo.2024.1449326. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 39286269 Free PMC article.
-
Vasopressin and oxytocin excite BNST neurons via oxytocin receptors, which reduce anxious arousal.Cell Rep. 2025 Jun 24;44(6):115768. doi: 10.1016/j.celrep.2025.115768. Epub 2025 Jun 3. Cell Rep. 2025. PMID: 40471787 Free PMC article.
-
Like sisters but not twins - vasopressin and oxytocin excite BNST neurons via cell type-specific expression of oxytocin receptor to reduce anxious arousal.bioRxiv [Preprint]. 2024 Sep 28:2024.09.06.611656. doi: 10.1101/2024.09.06.611656. bioRxiv. 2024. Update in: Cell Rep. 2025 Jun 24;44(6):115768. doi: 10.1016/j.celrep.2025.115768. PMID: 39282380 Free PMC article. Updated. Preprint.
References
-
- Alheid GF, 2003. Extended amygdala and basal forebrain. Ann. N. Y. Acad. Sci. 985, 185–205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
