

Structural Modeling of the Treponema pallidum Outer Membrane Protein Repertoire: a Road Map for Deconvolution of Syphilis Pathogenesis and Development of a Syphilis Vaccine
- PMID: 33972353
- PMCID: PMC8407342
- DOI: 10.1128/JB.00082-21
Structural Modeling of the Treponema pallidum Outer Membrane Protein Repertoire: a Road Map for Deconvolution of Syphilis Pathogenesis and Development of a Syphilis Vaccine
Abstract
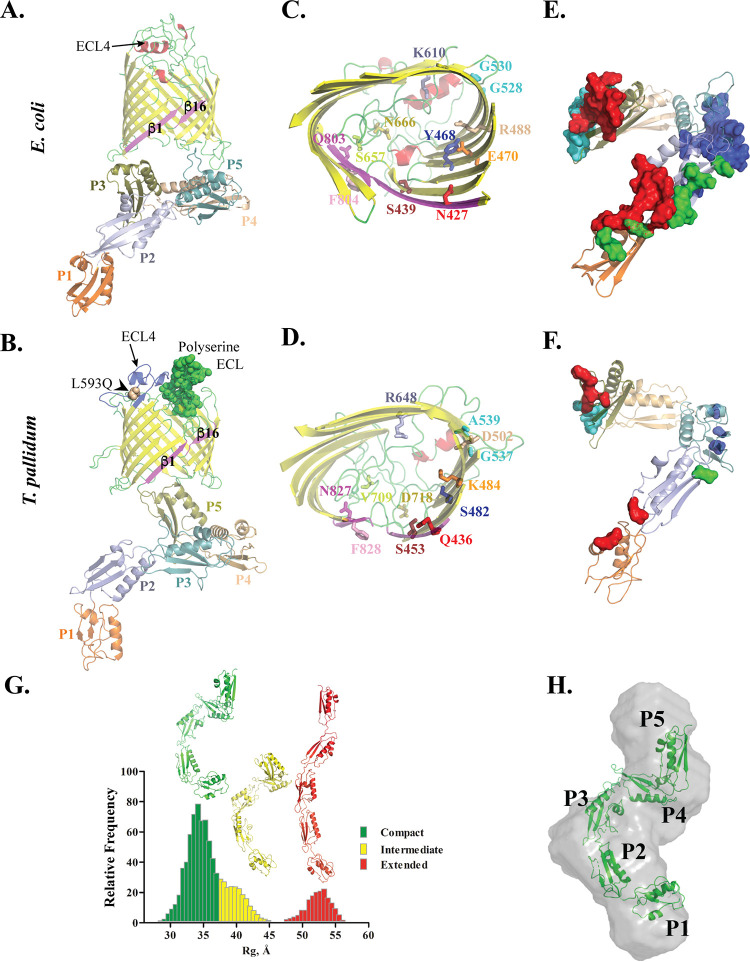
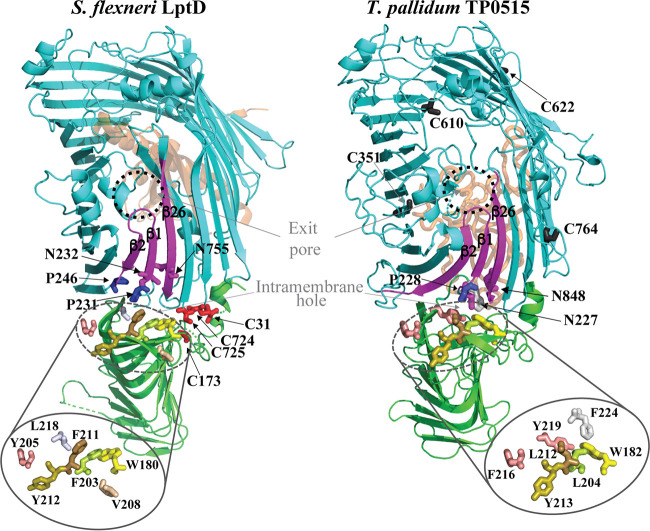
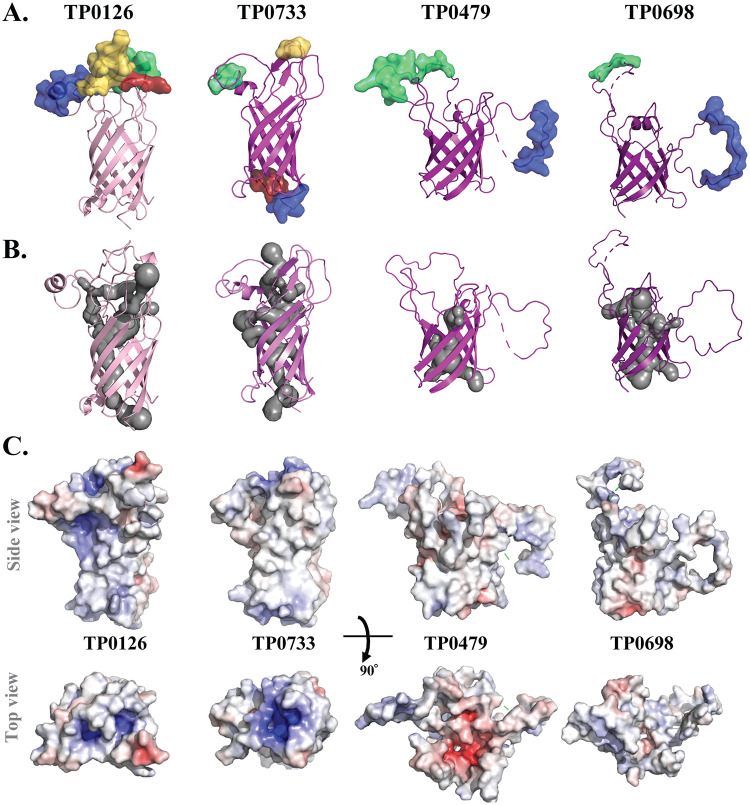
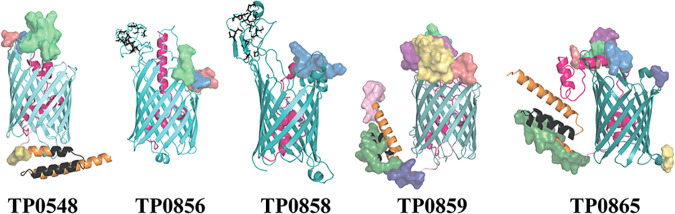
Treponema pallidum, an obligate human pathogen, has an outer membrane (OM) whose physical properties, ultrastructure, and composition differ markedly from those of phylogenetically distant Gram-negative bacteria. We developed structural models for the outer membrane protein (OMP) repertoire (OMPeome) of T. pallidum Nichols using solved Gram-negative structures, computational tools, and small-angle X-ray scattering (SAXS) of selected recombinant periplasmic domains. The T. pallidum "OMPeome" harbors two "stand-alone" proteins (BamA and LptD) involved in OM biogenesis and four paralogous families involved in the influx/efflux of small molecules: 8-stranded β-barrels, long-chain-fatty-acid transporters (FadLs), OM factors (OMFs) for efflux pumps, and T. pallidum repeat proteins (Tprs). BamA (TP0326), the central component of a β-barrel assembly machine (BAM)/translocation and assembly module (TAM) hybrid, possesses a highly flexible polypeptide-transport-associated (POTRA) 1-5 arm predicted to interact with TamB (TP0325). TP0515, an LptD ortholog, contains a novel, unstructured C-terminal domain that models inside the β-barrel. T. pallidum has four 8-stranded β-barrels, each containing positively charged extracellular loops that could contribute to pathogenesis. Three of five FadL-like orthologs have a novel α-helical, presumptively periplasmic C-terminal extension. SAXS and structural modeling further supported the bipartite membrane topology and tridomain architecture of full-length members of the Tpr family. T. pallidum's two efflux pumps presumably extrude noxious small molecules via four coexpressed OMFs with variably charged tunnels. For BamA, LptD, and OMFs, we modeled the molecular machines that deliver their substrates into the OM or external milieu. The spirochete's extended families of OM transporters collectively confer a broad capacity for nutrient uptake. The models also furnish a structural road map for vaccine development. IMPORTANCE The unusual outer membrane (OM) of T. pallidum, the syphilis spirochete, is the ultrastructural basis for its well-recognized capacity for invasiveness, immune evasion, and persistence. In recent years, we have made considerable progress in identifying T. pallidum's repertoire of OMPs. Here, we developed three-dimensional (3D) models for the T. pallidum Nichols OMPeome using structural modeling, bioinformatics, and solution scattering. The OM contains three families of OMP transporters, an OMP family involved in the extrusion of noxious molecules, and two "stand-alone" proteins involved in OM biogenesis. This work represents a major advance toward elucidating host-pathogen interactions during syphilis; understanding how T. pallidum, an extreme auxotroph, obtains a wide array of biomolecules from its obligate human host; and developing a vaccine with global efficacy.
Keywords: Treponema pallidum; bioinformatics; outer membrane proteins; structural biology; syphilis; vaccines.
Figures

Similar articles
-
Extracellular Loops of the Treponema pallidum FadL Orthologs TP0856 and TP0858 Elicit IgG Antibodies and IgG+-Specific B-Cells in the Rabbit Model of Experimental Syphilis.mBio. 2022 Aug 30;13(4):e0163922. doi: 10.1128/mbio.01639-22. Epub 2022 Jul 12. mBio. 2022. PMID: 35862766 Free PMC article.
-
A Homology Model Reveals Novel Structural Features and an Immunodominant Surface Loop/Opsonic Target in the Treponema pallidum BamA Ortholog TP_0326.J Bacteriol. 2015 Jun;197(11):1906-20. doi: 10.1128/JB.00086-15. Epub 2015 Mar 30. J Bacteriol. 2015. PMID: 25825429 Free PMC article.
-
Sequence Variation of Rare Outer Membrane Protein β-Barrel Domains in Clinical Strains Provides Insights into the Evolution of Treponema pallidum subsp. pallidum, the Syphilis Spirochete.mBio. 2018 Jun 12;9(3):e01006-18. doi: 10.1128/mBio.01006-18. mBio. 2018. PMID: 29895642 Free PMC article.
-
The Treponema pallidum Outer Membrane.Curr Top Microbiol Immunol. 2018;415:1-38. doi: 10.1007/82_2017_44. Curr Top Microbiol Immunol. 2018. PMID: 28849315 Free PMC article. Review.
-
Polypeptides of Treponema pallidum: progress toward understanding their structural, functional, and immunologic roles. Treponema Pallidum Polypeptide Research Group.Microbiol Rev. 1993 Sep;57(3):750-79. doi: 10.1128/mr.57.3.750-779.1993. Microbiol Rev. 1993. PMID: 8246847 Free PMC article. Review.
Cited by
-
Clinical Presentation of Early Syphilis and Genomic Sequences of Treponema pallidum Strains in Patient Specimens and Isolates Obtained by Rabbit Inoculation.J Infect Dis. 2024 Dec 16;230(6):e1322-e1333. doi: 10.1093/infdis/jiae322. J Infect Dis. 2024. PMID: 38884588 Free PMC article.
-
Full-length TprK of Treponema pallidum subsp. pallidum in lipid nanodiscs is a monomeric porin.Enzyme Microb Technol. 2022 Jan;153:109897. doi: 10.1016/j.enzmictec.2021.109897. Epub 2021 Aug 25. Enzyme Microb Technol. 2022. PMID: 34670182 Free PMC article.
-
Use of Epivolve phage display to generate a monoclonal antibody with opsonic activity directed against a subdominant epitope on extracellular loop 4 of Treponema pallidum BamA (TP0326).Front Immunol. 2023 Aug 22;14:1222267. doi: 10.3389/fimmu.2023.1222267. eCollection 2023. Front Immunol. 2023. PMID: 37675118 Free PMC article.
-
Proteomic analysis of the Treponema pallidum subsp. pallidum SS14 strain: coverage and comparison with the Nichols strain proteome.Front Microbiol. 2024 Dec 11;15:1505893. doi: 10.3389/fmicb.2024.1505893. eCollection 2024. Front Microbiol. 2024. PMID: 39723147 Free PMC article.
-
Extracellular Loops of the Treponema pallidum FadL Orthologs TP0856 and TP0858 Elicit IgG Antibodies and IgG+-Specific B-Cells in the Rabbit Model of Experimental Syphilis.mBio. 2022 Aug 30;13(4):e0163922. doi: 10.1128/mbio.01639-22. Epub 2022 Jul 12. mBio. 2022. PMID: 35862766 Free PMC article.
References
-
- Radolf JD, Tramont EC, Salazar JC. 2019. Syphilis (Treponema pallidum), p 2865–2892. In Bennett JE, Dolin R, Blaser MJ (ed), Mandell, Douglas, and Bennett’s principles and practice of infectious diseases, 9th ed. Elsevier, Philadelphia, PA.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
