

Inner hair cell stereocilia are embedded in the tectorial membrane
- PMID: 33972539
- PMCID: PMC8110531
- DOI: 10.1038/s41467-021-22870-1
Inner hair cell stereocilia are embedded in the tectorial membrane
Abstract
Mammalian hearing depends on sound-evoked displacements of the stereocilia of inner hair cells (IHCs), which cause the endogenous mechanoelectrical transducer channels to conduct inward currents of cations including Ca2+. Due to their presumed lack of contacts with the overlaying tectorial membrane (TM), the putative stimulation mechanism for these stereocilia is by means of the viscous drag of the surrounding endolymph. However, despite numerous efforts to characterize the TM by electron microscopy and other techniques, the exact IHC stereocilia-TM relationship remains elusive. Here we show that Ca2+-rich filamentous structures, that we call Ca2+ ducts, connect the TM to the IHC stereocilia to enable mechanical stimulation by the TM while also ensuring the stereocilia access to TM Ca2+. Our results call for a reassessment of the stimulation mechanism for the IHC stereocilia and the TM role in hearing.
Conflict of interest statement
The authors declare no competing interests.
Figures
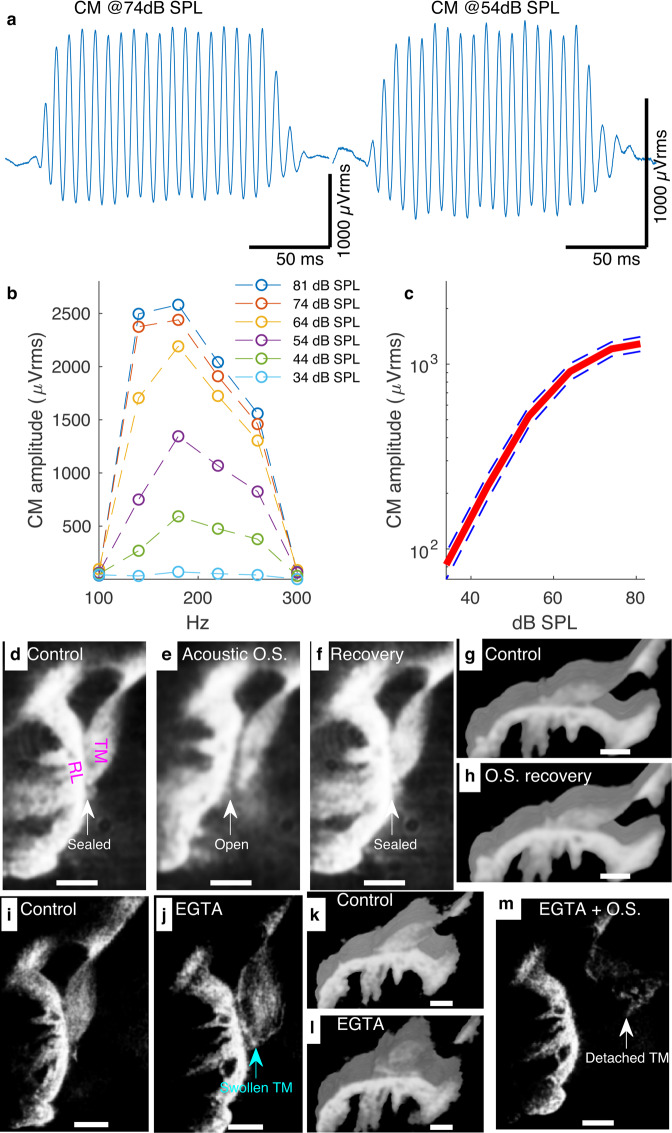
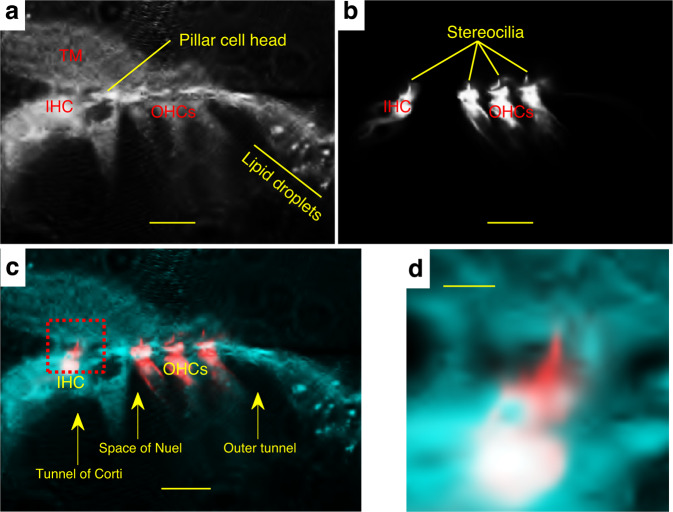
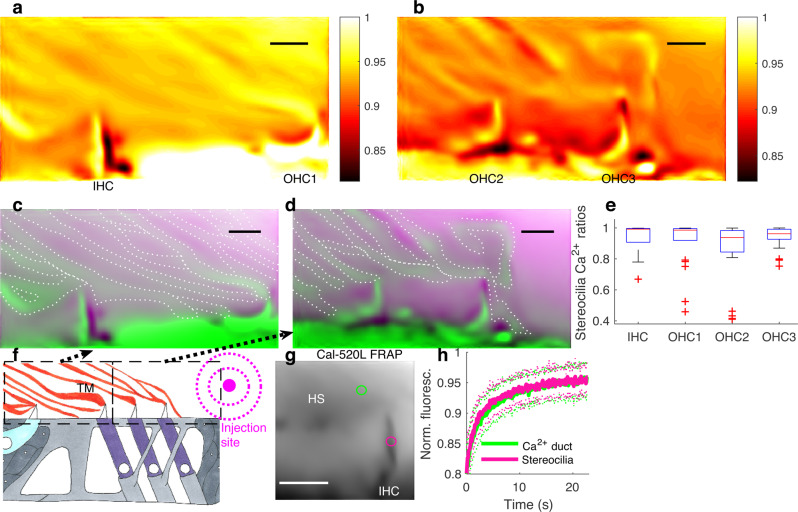
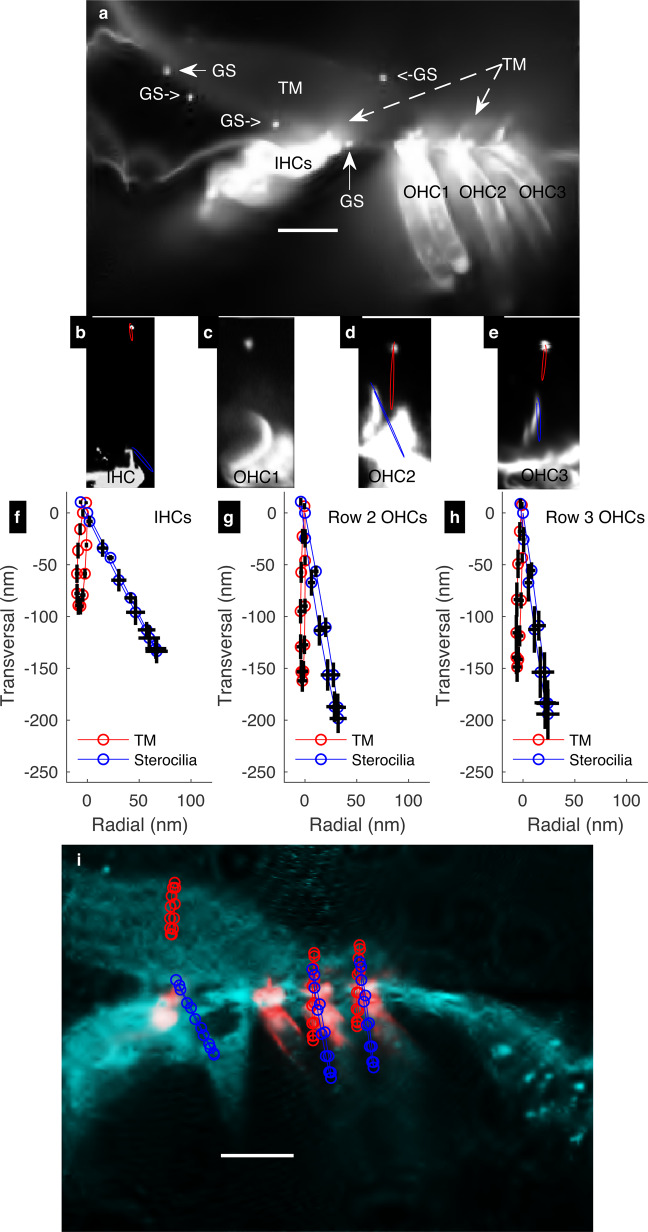
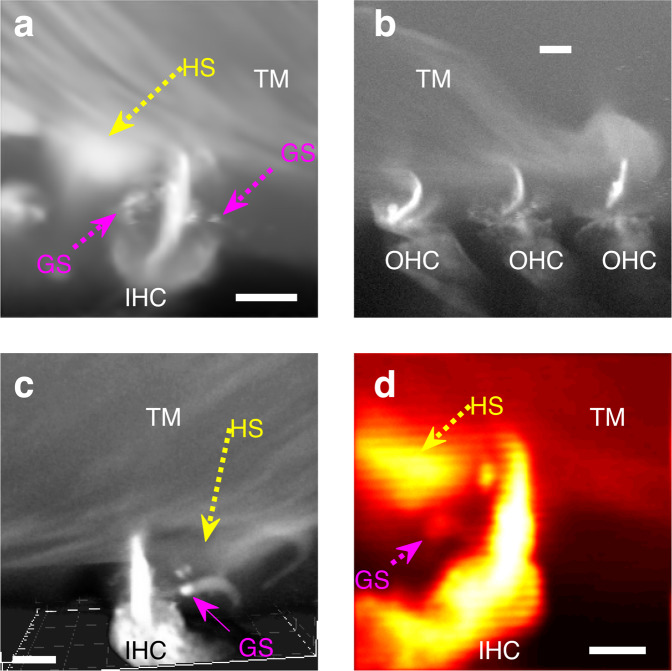
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
