

A Ran-binding protein facilitates nuclear import of human papillomavirus type 16
- PMID: 33974675
- PMCID: PMC8139508
- DOI: 10.1371/journal.ppat.1009580
A Ran-binding protein facilitates nuclear import of human papillomavirus type 16
Abstract
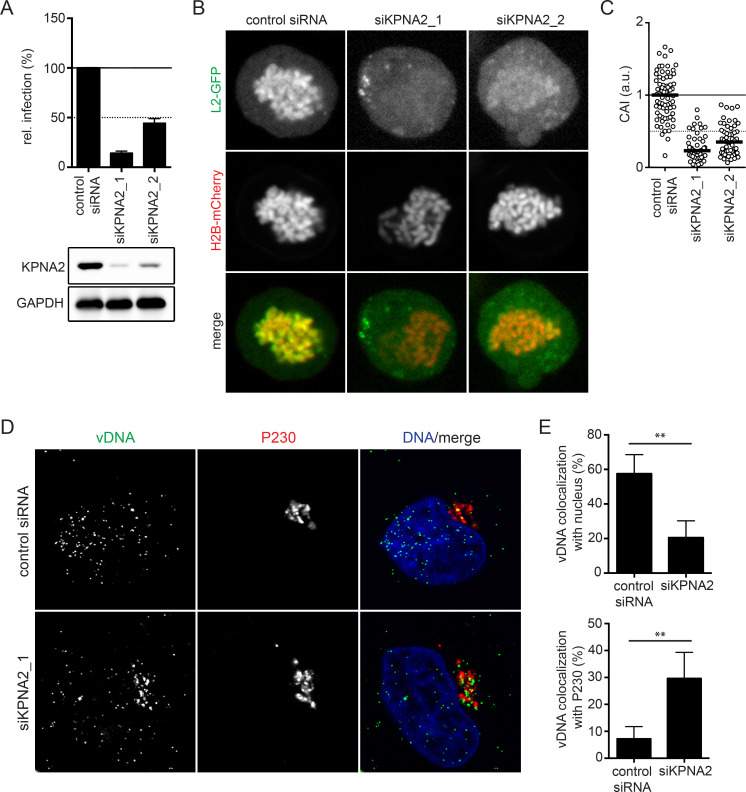
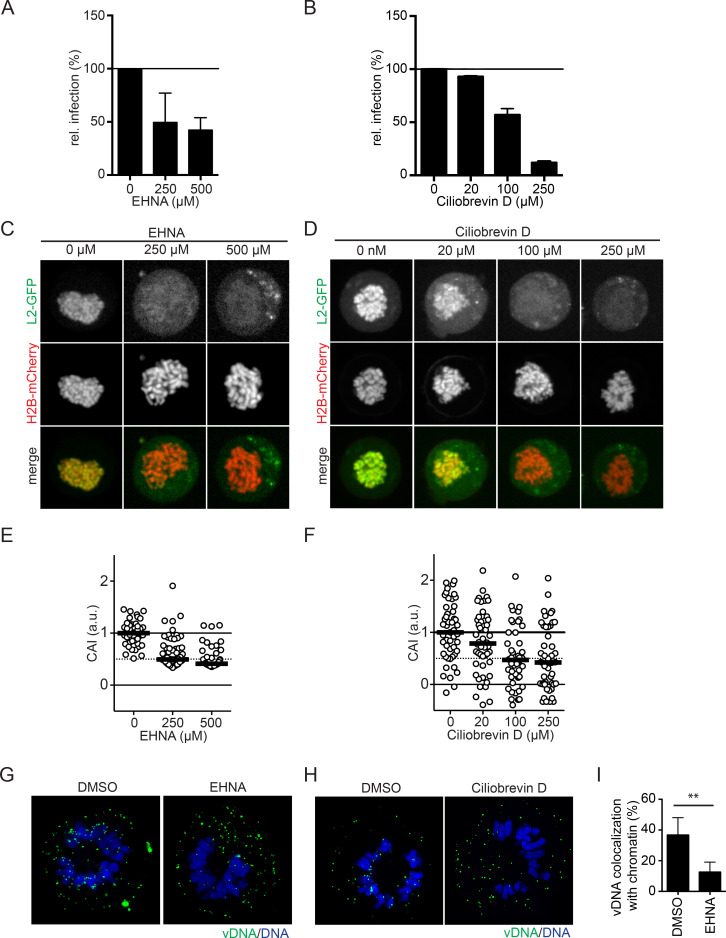
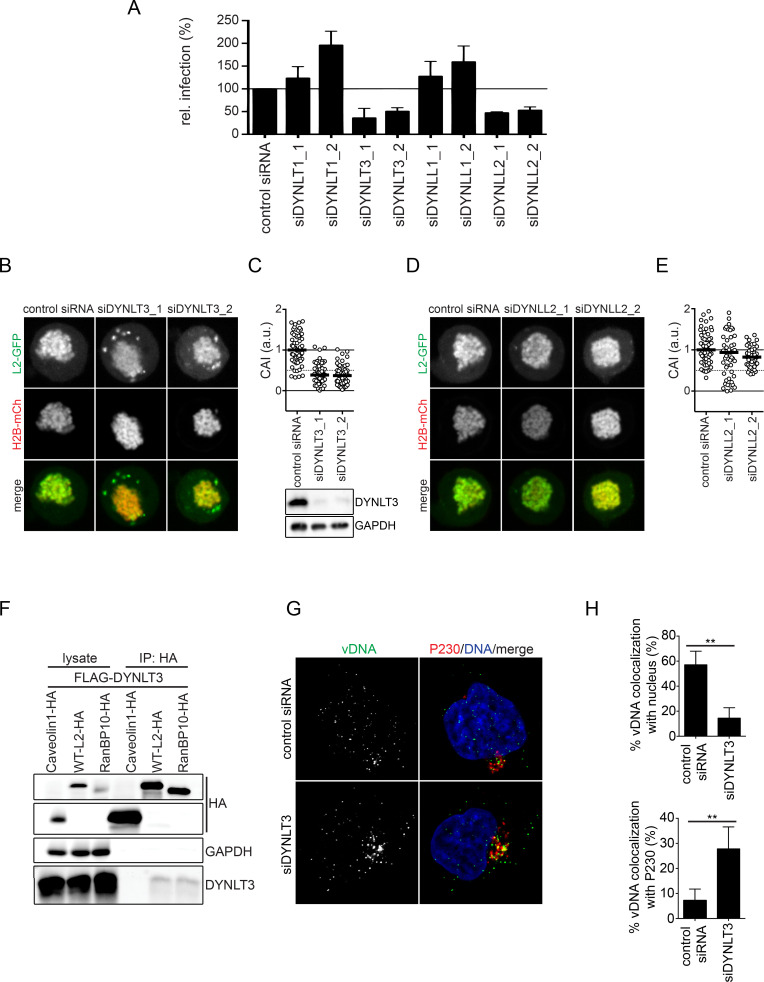
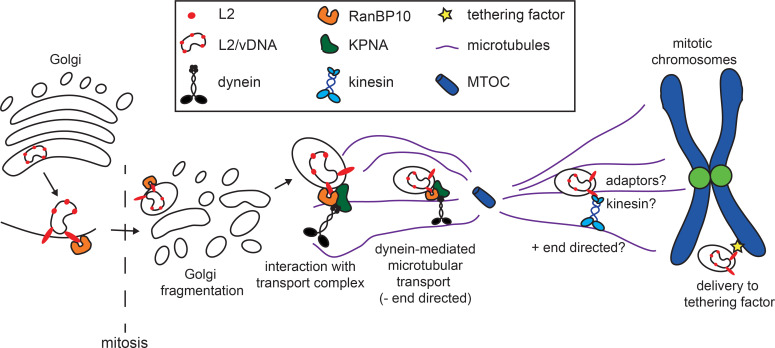
Human papillomaviruses (HPVs) utilize an atypical mode of nuclear import during cell entry. Residing in the Golgi apparatus until mitosis onset, a subviral complex composed of the minor capsid protein L2 and viral DNA (L2/vDNA) is imported into the nucleus after nuclear envelope breakdown by associating with mitotic chromatin. In this complex, L2 plays a crucial role in the interactions with cellular factors that enable delivery and ultimately tethering of the viral genome to mitotic chromatin. To date, the cellular proteins facilitating these steps remain unknown. Here, we addressed which cellular proteins may be required for this process. Using label-free mass spectrometry, biochemical assays, microscopy, and functional virological assays, we discovered that L2 engages a hitherto unknown protein complex of Ran-binding protein 10 (RanBP10), karyopherin alpha2 (KPNA2), and dynein light chain DYNLT3 to facilitate transport towards mitotic chromatin. Thus, our study not only identifies novel cellular interactors and mechanism that facilitate a poorly understood step in HPV entry, but also a novel cellular transport complex.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
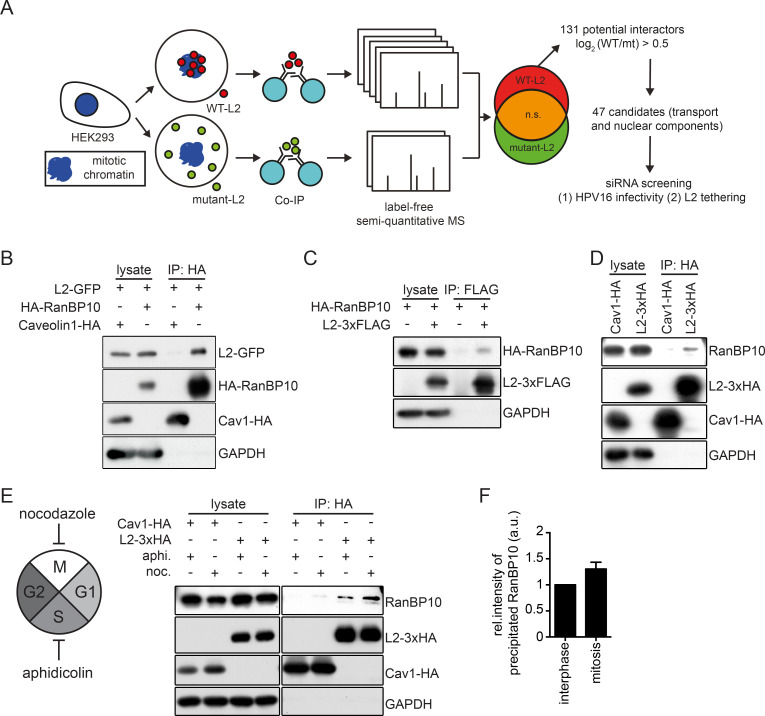
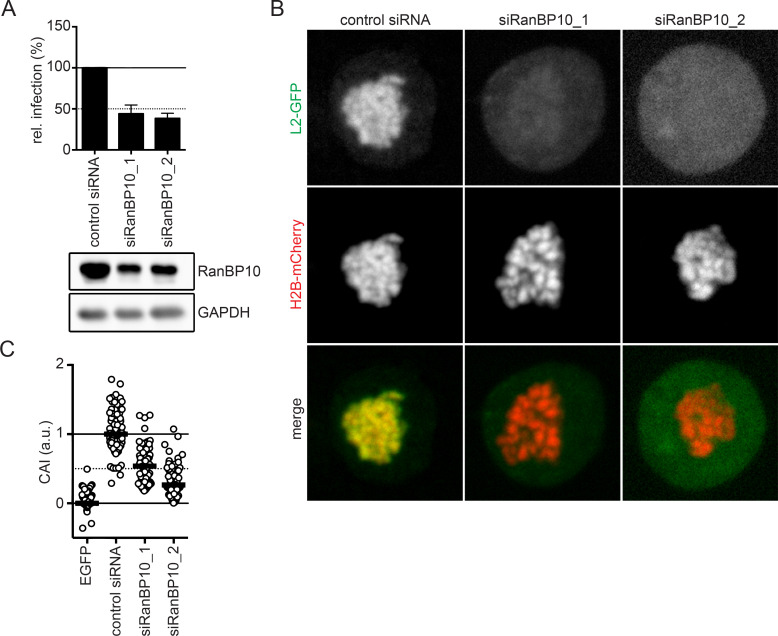
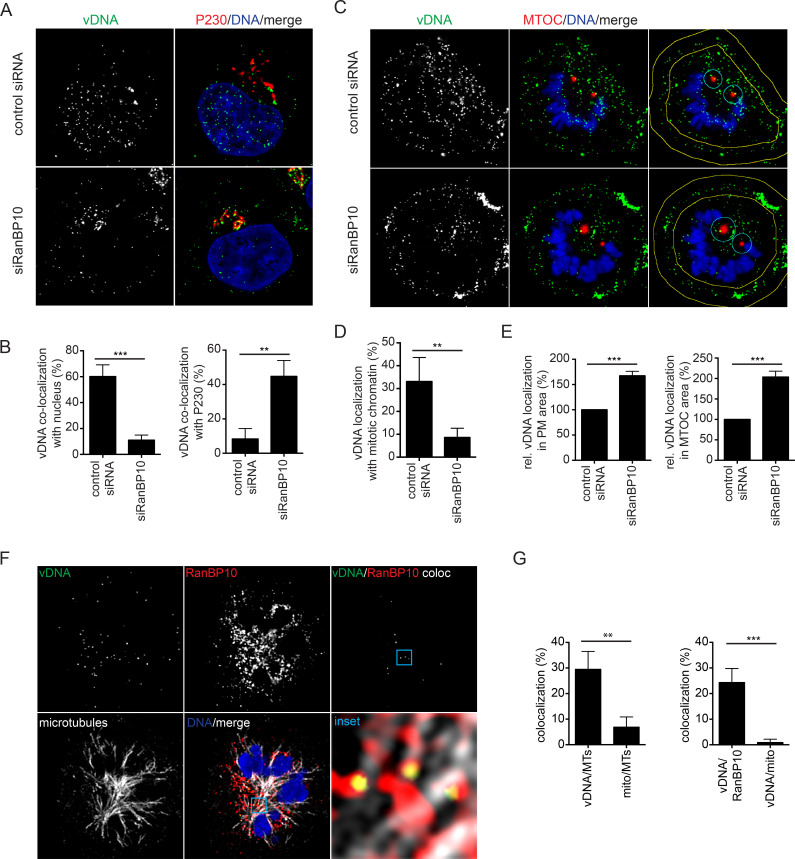
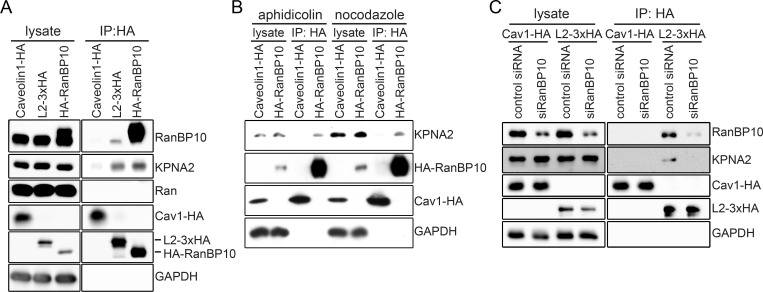
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
