

Estrogens and the circadian system
- PMID: 33975754
- PMCID: PMC8573061
- DOI: 10.1016/j.semcdb.2021.04.010
Estrogens and the circadian system
Abstract
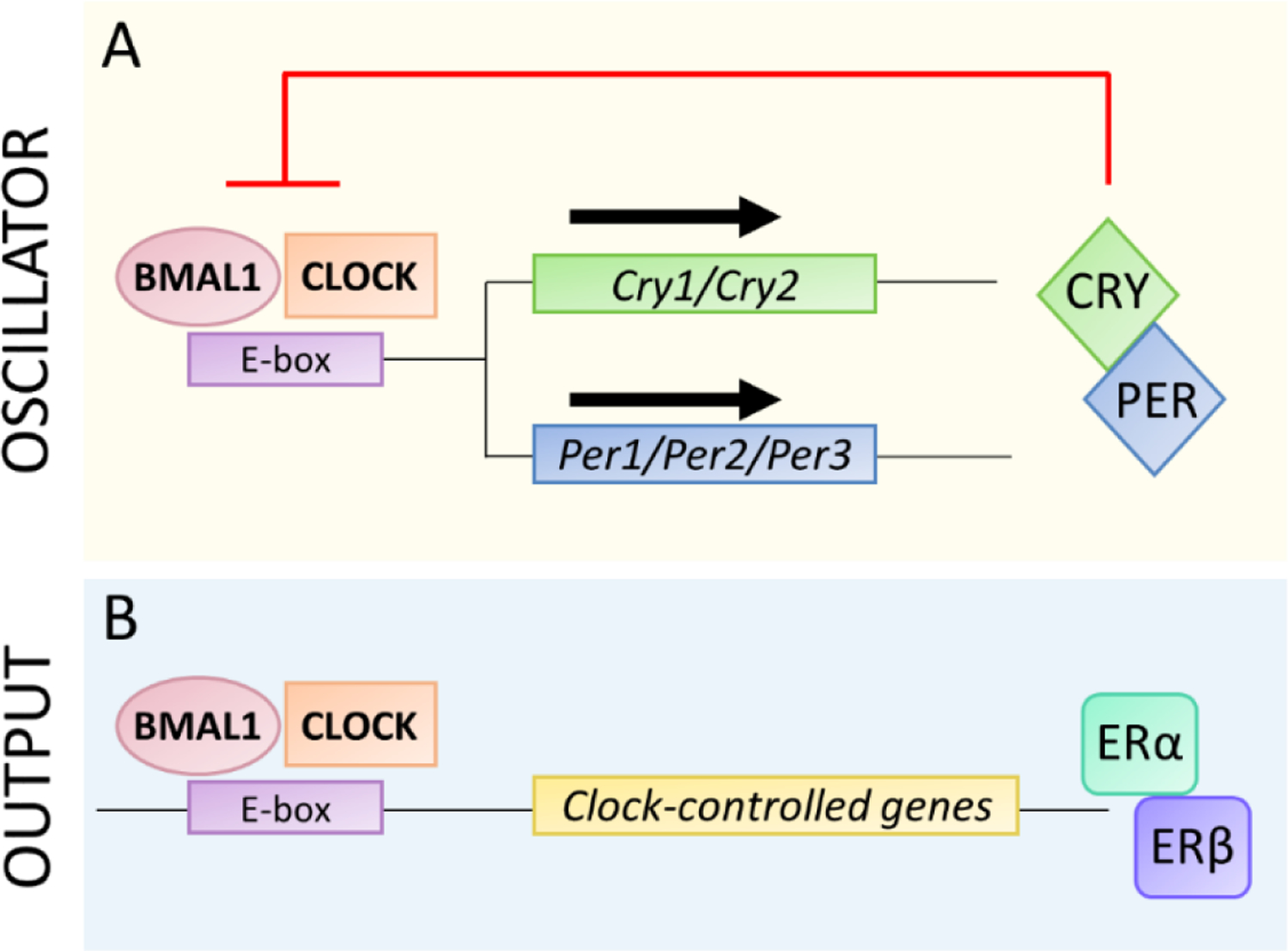
Circadian rhythms are ~24 h cycles of behavior and physiology that are generated by a network of molecular clocks located in nearly every tissue in the body. In mammals, the circadian system is organized hierarchically such that the suprachiasmatic nucleus (SCN) is the main circadian clock that receives light information from the eye and entrains to the light-dark cycle. The SCN then coordinates the timing of tissue clocks so internal rhythms are aligned with environmental cycles. Estrogens interact with the circadian system to regulate biological processes. At the molecular level, estrogens and circadian genes interact to regulate gene expression and cell biology. Estrogens also regulate circadian behavior across the estrous cycle. The timing of ovulation during the estrous cycle requires coincident estrogen and SCN signals. Studies using circadian gene reporter mice have also elucidated estrogen regulation of peripheral tissue clocks and metabolic rhythms. This review synthesizes current understanding of the interplay between estrogens and the circadian system, with a focus on female rodents, in regulating molecular, physiological, and behavioral processes.
Keywords: Circadian rhythm; Clock genes; Estradiol; Female; Mouse; Suprachiasmatic nucleus.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures




References
-
- Halberg F, [Physiologic 24-hour periodicity; general and procedural considerations with reference to the adrenal cycle], Int Z Vitaminforsch Beih 10 (1959) 225–96. - PubMed
-
- Pittendrigh CS, Daan S, A functional analysis of circadian pacemakers in nocturnal rodents. I. The stability and lability of spontaneous frequency., J Comp Physiol 106 (1976) 333–55.
-
- DeCoursey PJ, Krulas JR, Behavior of SCN-lesioned chipmunks in natural habitat: a pilot study, J Biol Rhythms 13(3) (1998) 229–44. - PubMed
-
- Hurd MW, Ralph MR, The significance of circadian organization for longevity in the golden hamster, J Biol Rhythms 13(5) (1998) 430–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
