

Expression of FoxP2 in the basal ganglia regulates vocal motor sequences in the adult songbird
- PMID: 33976169
- PMCID: PMC8113549
- DOI: 10.1038/s41467-021-22918-2
Expression of FoxP2 in the basal ganglia regulates vocal motor sequences in the adult songbird
Abstract
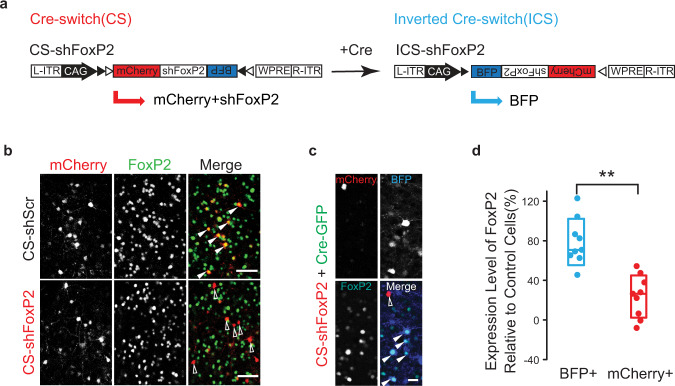
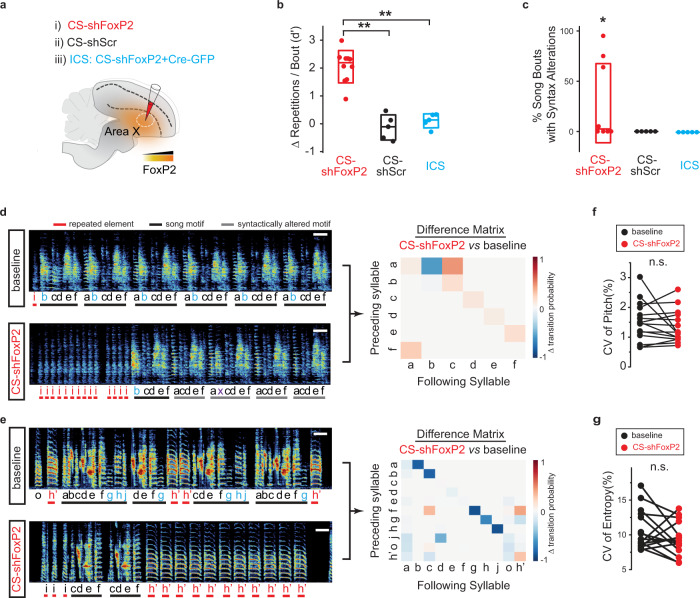
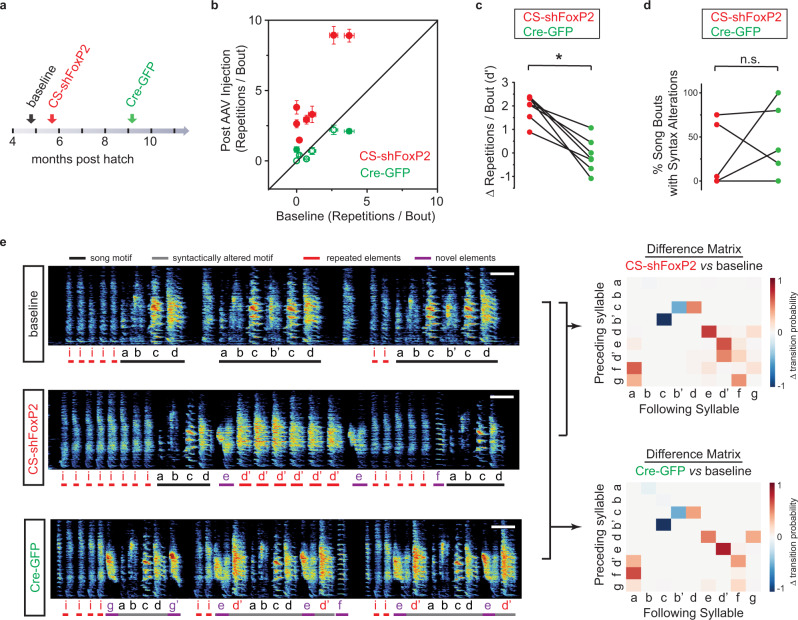
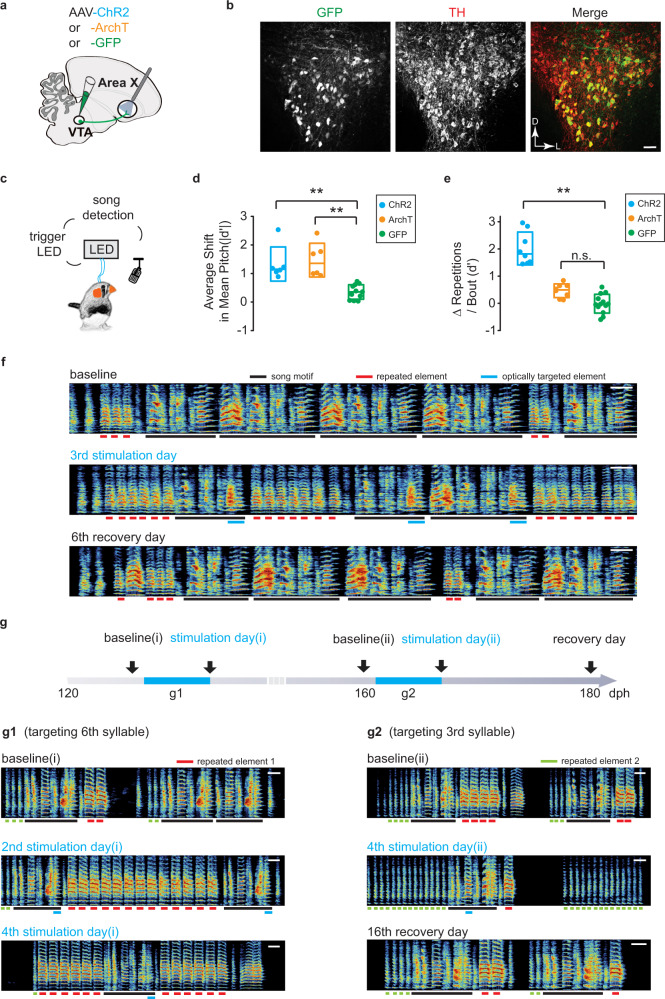
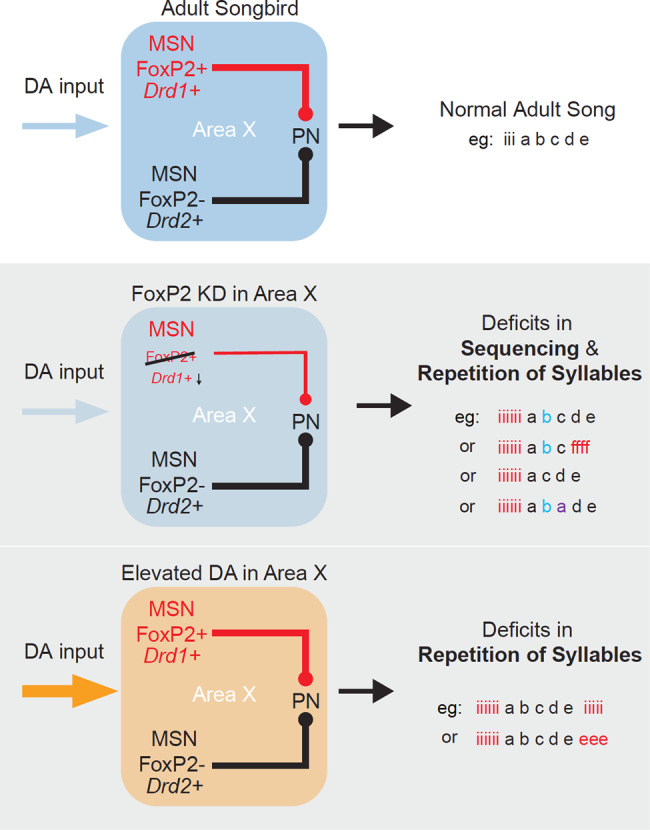
Disruption of the transcription factor FoxP2, which is enriched in the basal ganglia, impairs vocal development in humans and songbirds. The basal ganglia are important for the selection and sequencing of motor actions, but the circuit mechanisms governing accurate sequencing of learned vocalizations are unknown. Here, we show that expression of FoxP2 in the basal ganglia is vital for the fluent initiation and termination of birdsong, as well as the maintenance of song syllable sequencing in adulthood. Knockdown of FoxP2 imbalances dopamine receptor expression across striatal direct-like and indirect-like pathways, suggesting a role of dopaminergic signaling in regulating vocal motor sequencing. Confirming this prediction, we show that phasic dopamine activation, and not inhibition, during singing drives repetition of song syllables, thus also impairing fluent initiation and termination of birdsong. These findings demonstrate discrete circuit origins for the dysfluent repetition of vocal elements in songbirds, with implications for speech disorders.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Incomplete and inaccurate vocal imitation after knockdown of FoxP2 in songbird basal ganglia nucleus Area X.PLoS Biol. 2007 Dec;5(12):e321. doi: 10.1371/journal.pbio.0050321. PLoS Biol. 2007. PMID: 18052609 Free PMC article.
-
Beyond Critical Period Learning: Striatal FoxP2 Affects the Active Maintenance of Learned Vocalizations in Adulthood.eNeuro. 2019 Apr 15;6(2):ENEURO.0071-19.2019. doi: 10.1523/ENEURO.0071-19.2019. eCollection 2019 Mar-Apr. eNeuro. 2019. PMID: 31001575 Free PMC article.
-
Expression analysis of the speech-related genes FoxP1 and FoxP2 and their relation to singing behavior in two songbird species.J Exp Biol. 2013 Oct 1;216(Pt 19):3682-92. doi: 10.1242/jeb.085886. J Exp Biol. 2013. PMID: 24006346 Free PMC article.
-
From songbird to humans: The multifaceted roles of FOXP2 in speech and motor learning.Neurosci Biobehav Rev. 2024 Dec;167:105936. doi: 10.1016/j.neubiorev.2024.105936. Epub 2024 Nov 6. Neurosci Biobehav Rev. 2024. PMID: 39510218 Review.
-
Twitter evolution: converging mechanisms in birdsong and human speech.Nat Rev Neurosci. 2010 Nov;11(11):747-59. doi: 10.1038/nrn2931. Nat Rev Neurosci. 2010. PMID: 20959859 Review.
Cited by
-
Cellular transcriptomics reveals evolutionary identities of songbird vocal circuits.Science. 2021 Feb 12;371(6530):eabd9704. doi: 10.1126/science.abd9704. Science. 2021. PMID: 33574185 Free PMC article.
-
Single-cell transcriptomics reveals the brain evolution of web-building spiders.Nat Ecol Evol. 2023 Dec;7(12):2125-2142. doi: 10.1038/s41559-023-02238-y. Epub 2023 Nov 2. Nat Ecol Evol. 2023. PMID: 37919396 Free PMC article.
-
Molecular networks of the FOXP2 transcription factor in the brain.EMBO Rep. 2021 Aug 4;22(8):e52803. doi: 10.15252/embr.202152803. Epub 2021 Jul 14. EMBO Rep. 2021. PMID: 34260143 Free PMC article. Review.
-
What Is the Role of Thalamostriatal Circuits in Learning Vocal Sequences?Front Neural Circuits. 2021 Sep 22;15:724858. doi: 10.3389/fncir.2021.724858. eCollection 2021. Front Neural Circuits. 2021. PMID: 34630047 Free PMC article.
-
Wings of Change: aPKC/FoxP-dependent plasticity in steering motor neurons underlies operant self-learning in Drosophila.F1000Res. 2024 Jun 11;13:116. doi: 10.12688/f1000research.146347.2. eCollection 2024. F1000Res. 2024. PMID: 38779314 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
