

Bidirectional propagation of low frequency oscillations over the human hippocampal surface
- PMID: 33980852
- PMCID: PMC8115072
- DOI: 10.1038/s41467-021-22850-5
Bidirectional propagation of low frequency oscillations over the human hippocampal surface
Abstract
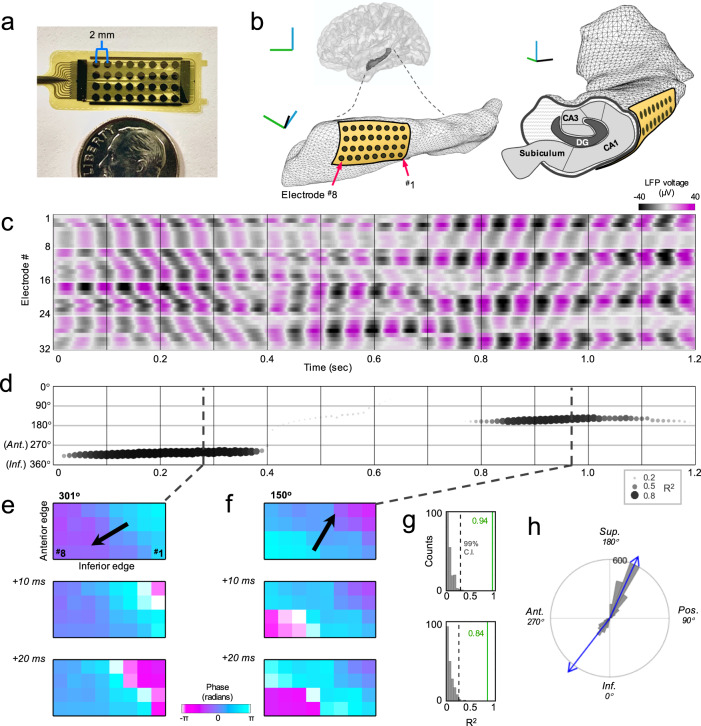
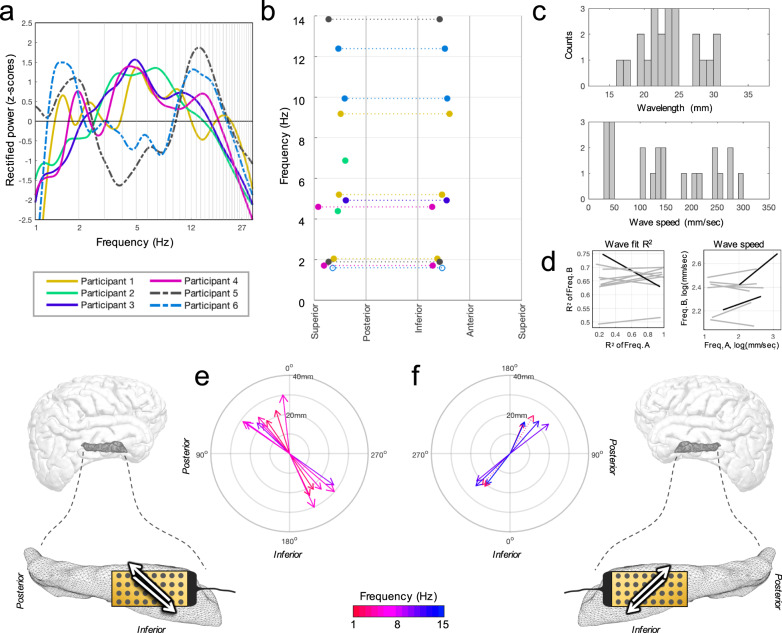
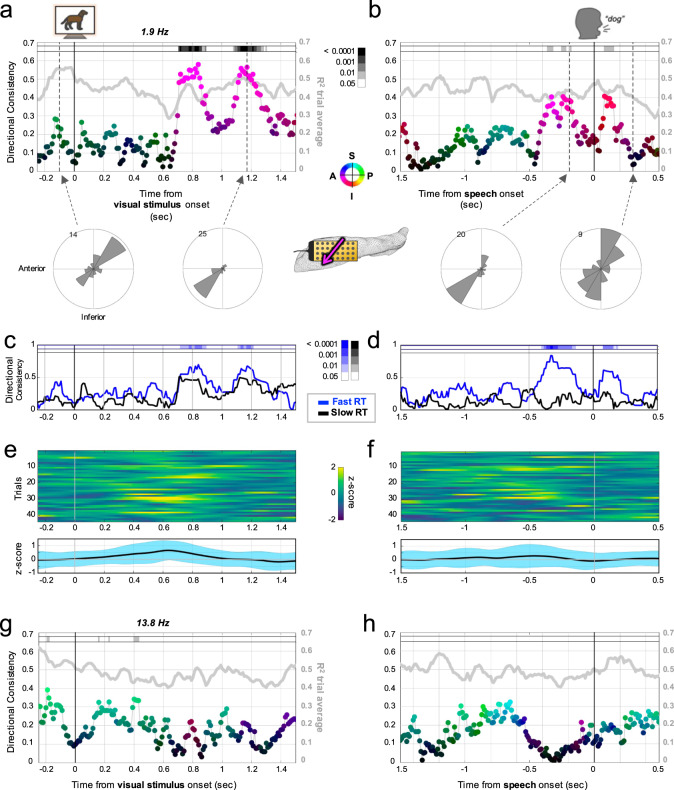
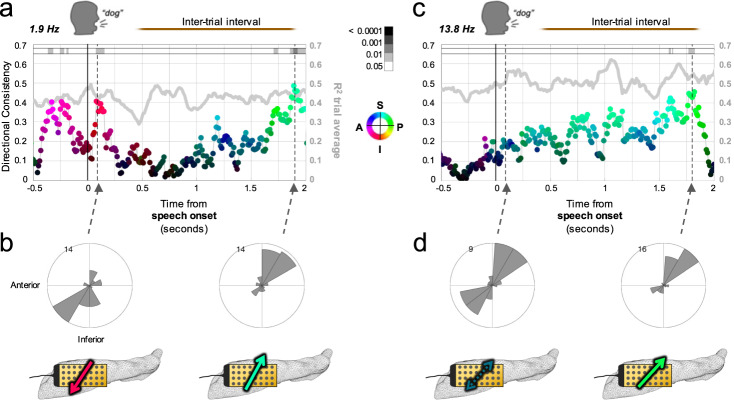
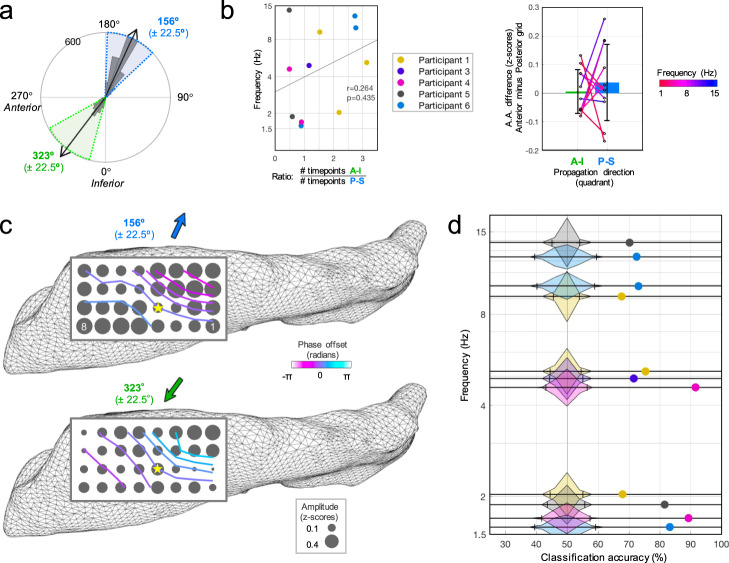
The hippocampus is diversely interconnected with other brain systems along its axis. Cycles of theta-frequency activity are believed to propagate from the septal to temporal pole, yet it is unclear how this one-way route supports the flexible cognitive capacities of this structure. We leveraged novel thin-film microgrid arrays conformed to the human hippocampal surface to track neural activity two-dimensionally in vivo. All oscillation frequencies identified between 1-15 Hz propagated across the tissue. Moreover, they dynamically shifted between two roughly opposite directions oblique to the long axis. This predominant propagation axis was mirrored across participants, hemispheres, and consciousness states. Directionality was modulated in a participant who performed a behavioral task, and it could be predicted by wave amplitude topography over the hippocampal surface. Our results show that propagation directions may thus represent distinct meso-scale network computations, operating along versatile spatiotemporal processing routes across the hippocampal body.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
