

Mechanical plasticity of collagen directs branch elongation in human mammary gland organoids
- PMID: 33980857
- PMCID: PMC8115695
- DOI: 10.1038/s41467-021-22988-2
Mechanical plasticity of collagen directs branch elongation in human mammary gland organoids
Abstract
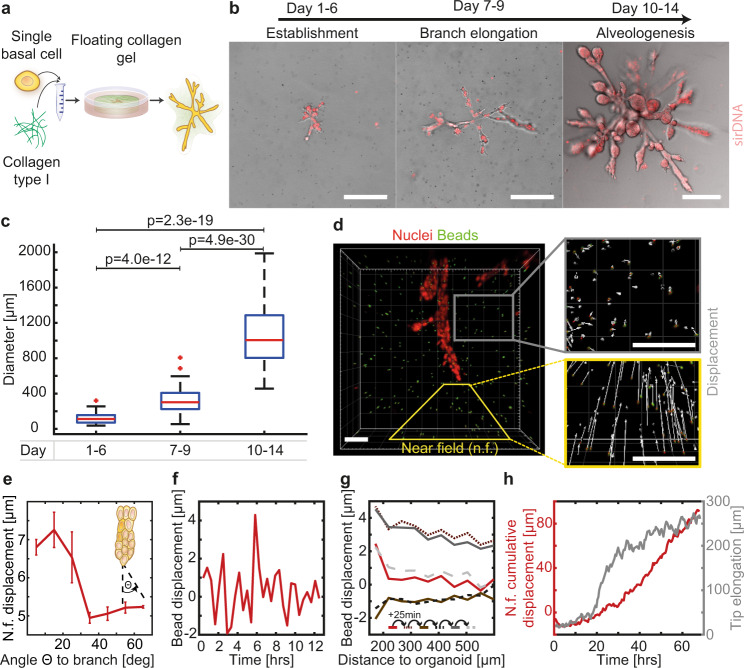
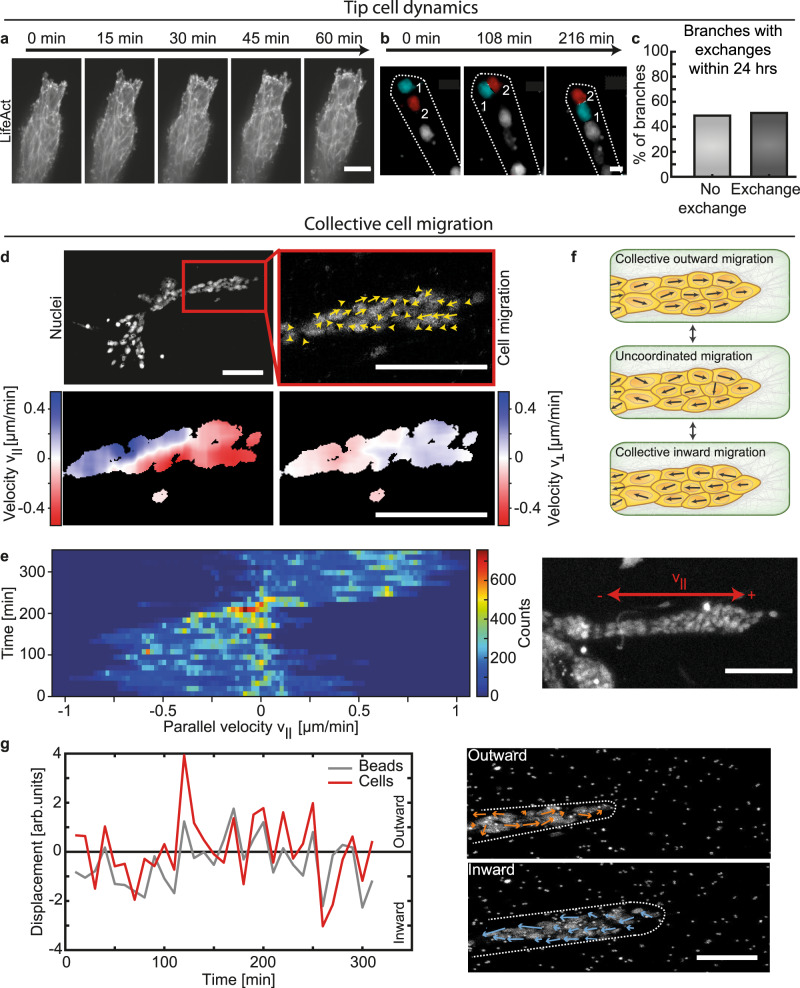
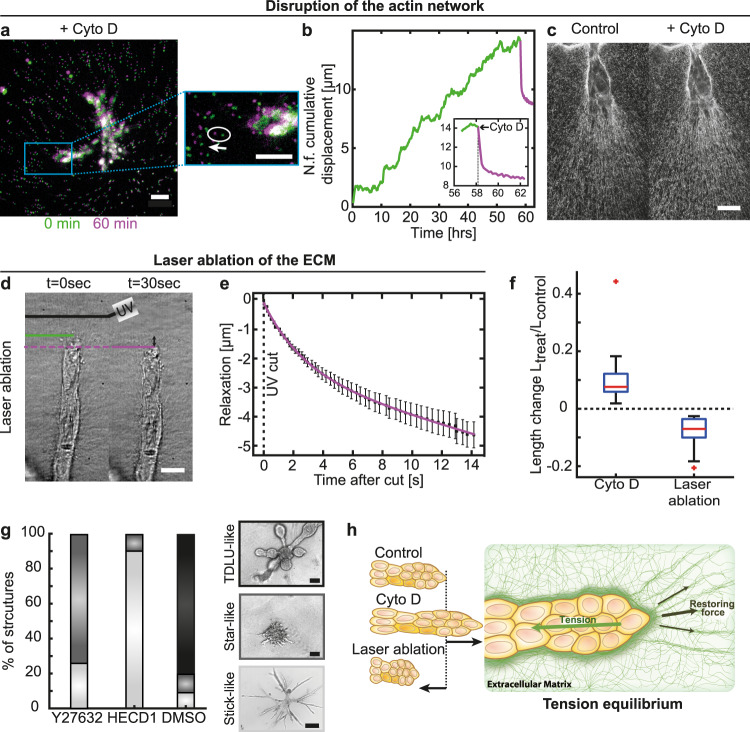
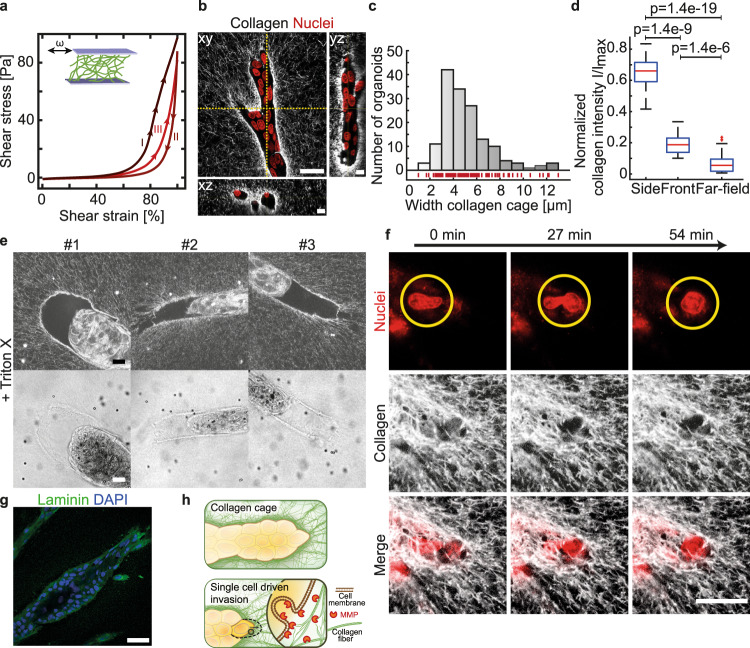
Epithelial branch elongation is a central developmental process during branching morphogenesis in diverse organs. This fundamental growth process into large arborized epithelial networks is accompanied by structural reorganization of the surrounding extracellular matrix (ECM), well beyond its mechanical linear response regime. Here, we report that epithelial ductal elongation within human mammary organoid branches relies on the non-linear and plastic mechanical response of the surrounding collagen. Specifically, we demonstrate that collective back-and-forth motion of cells within the branches generates tension that is strong enough to induce a plastic reorganization of the surrounding collagen network which results in the formation of mechanically stable collagen cages. Such matrix encasing in turn directs further tension generation, branch outgrowth and plastic deformation of the matrix. The identified mechanical tension equilibrium sets a framework to understand how mechanical cues can direct ductal branch elongation.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
From single cells to tissues: interactions between the matrix and human breast cells in real time.PLoS One. 2014 Apr 1;9(4):e93325. doi: 10.1371/journal.pone.0093325. eCollection 2014. PLoS One. 2014. PMID: 24691468 Free PMC article.
-
A Mammary Organoid Model to Study Branching Morphogenesis.Front Physiol. 2022 Mar 16;13:826107. doi: 10.3389/fphys.2022.826107. eCollection 2022. Front Physiol. 2022. PMID: 35399282 Free PMC article.
-
Mammary ductal elongation and myoepithelial migration are regulated by the composition of the extracellular matrix.J Microsc. 2013 Sep;251(3):212-23. doi: 10.1111/jmi.12017. Epub 2013 Feb 22. J Microsc. 2013. PMID: 23432616 Free PMC article.
-
3D culture models for studying branching morphogenesis in the mammary gland and mammalian lung.Biomaterials. 2019 Apr;198:135-145. doi: 10.1016/j.biomaterials.2018.08.043. Epub 2018 Aug 23. Biomaterials. 2019. PMID: 30174198 Free PMC article. Review.
-
Cellular foundations of mammary tubulogenesis.Semin Cell Dev Biol. 2014 Jul;31:124-31. doi: 10.1016/j.semcdb.2014.04.019. Epub 2014 Apr 18. Semin Cell Dev Biol. 2014. PMID: 24747369 Free PMC article. Review.
Cited by
-
Clinical applications of 3D normal and breast cancer organoids: A review of concepts and methods.Exp Biol Med (Maywood). 2022 Dec;247(24):2176-2183. doi: 10.1177/15353702221131877. Epub 2022 Nov 19. Exp Biol Med (Maywood). 2022. PMID: 36408534 Free PMC article. Review.
-
How to Study the Mechanobiology of Intestinal Epithelial Organoids? A Review of Culture Supports, Imaging Techniques, and Analysis Methods.Biol Cell. 2025 Apr;117(4):e70003. doi: 10.1111/boc.70003. Biol Cell. 2025. PMID: 40223609 Free PMC article. Review.
-
Mechanobiological engineering strategies for organoid culture.APL Bioeng. 2025 Jul 18;9(3):031501. doi: 10.1063/5.0275439. eCollection 2025 Sep. APL Bioeng. 2025. PMID: 40688244 Free PMC article. Review.
-
Organoid-on-a-chip: Current challenges, trends, and future scope toward medicine.Biomicrofluidics. 2023 Oct 27;17(5):051505. doi: 10.1063/5.0171350. eCollection 2023 Sep. Biomicrofluidics. 2023. PMID: 37900053 Free PMC article. Review.
-
Vascularized Tissue Organoids.Bioengineering (Basel). 2023 Jan 17;10(2):124. doi: 10.3390/bioengineering10020124. Bioengineering (Basel). 2023. PMID: 36829618 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
