

Dietary cis-9, trans-11-conjugated linoleic acid reduces amyloid β-protein accumulation and upregulates anti-inflammatory cytokines in an Alzheimer's disease mouse model
- PMID: 33980877
- PMCID: PMC8115273
- DOI: 10.1038/s41598-021-88870-9
Dietary cis-9, trans-11-conjugated linoleic acid reduces amyloid β-protein accumulation and upregulates anti-inflammatory cytokines in an Alzheimer's disease mouse model
Abstract
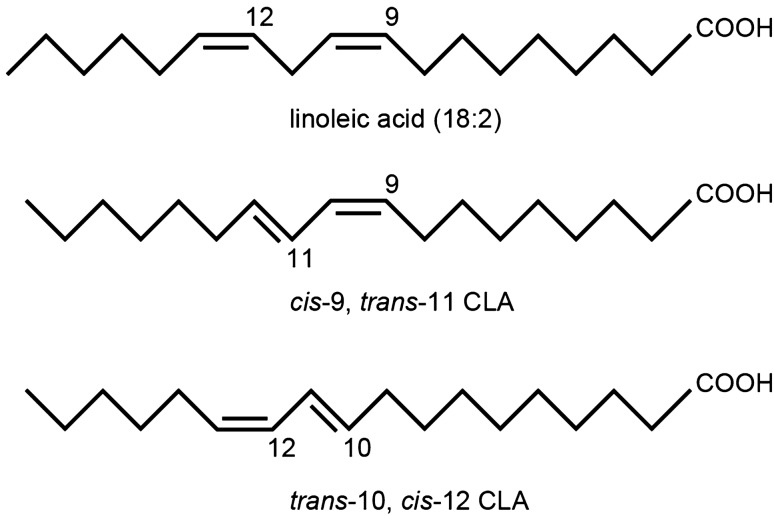
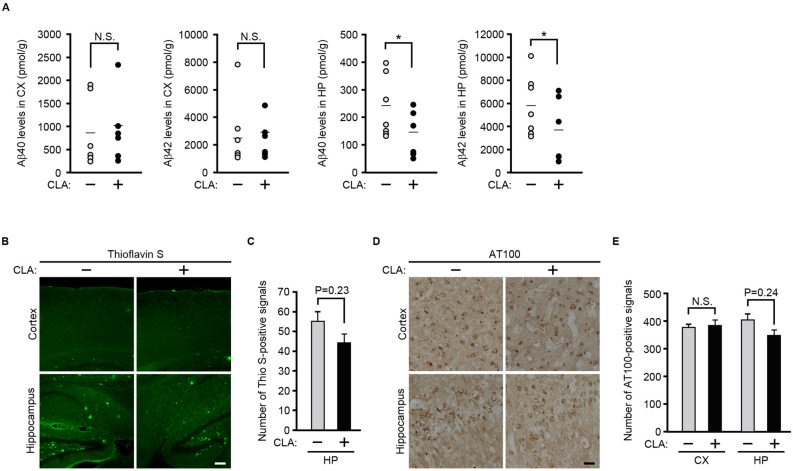
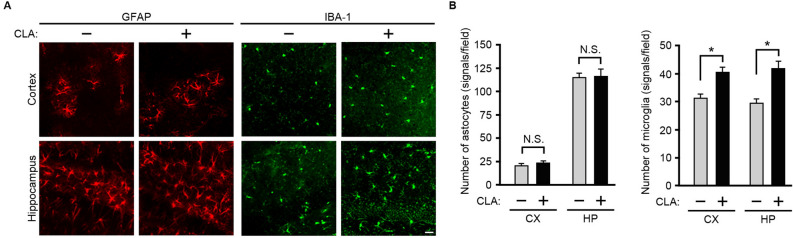
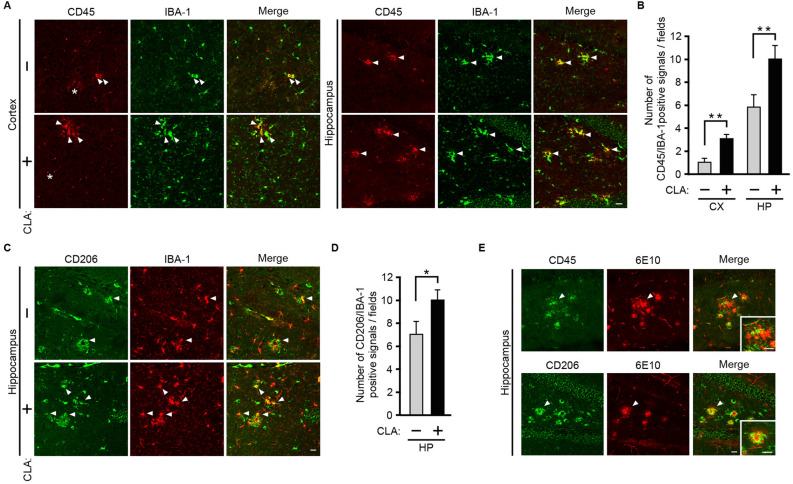
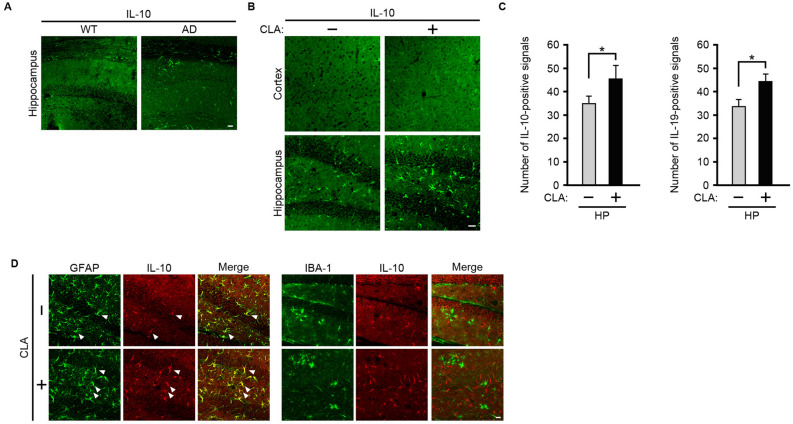
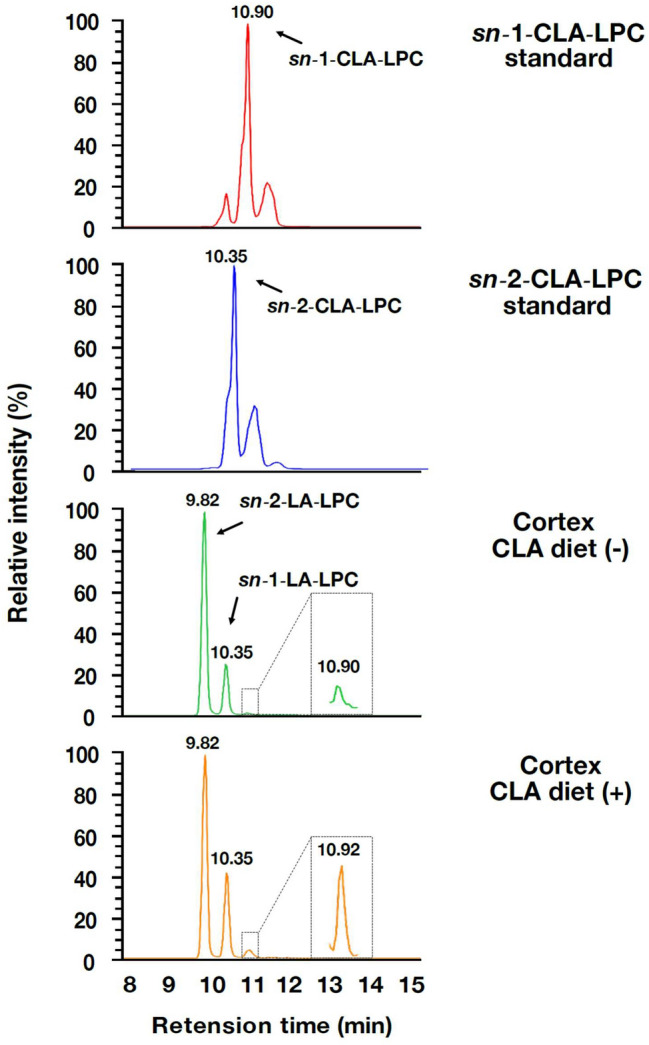
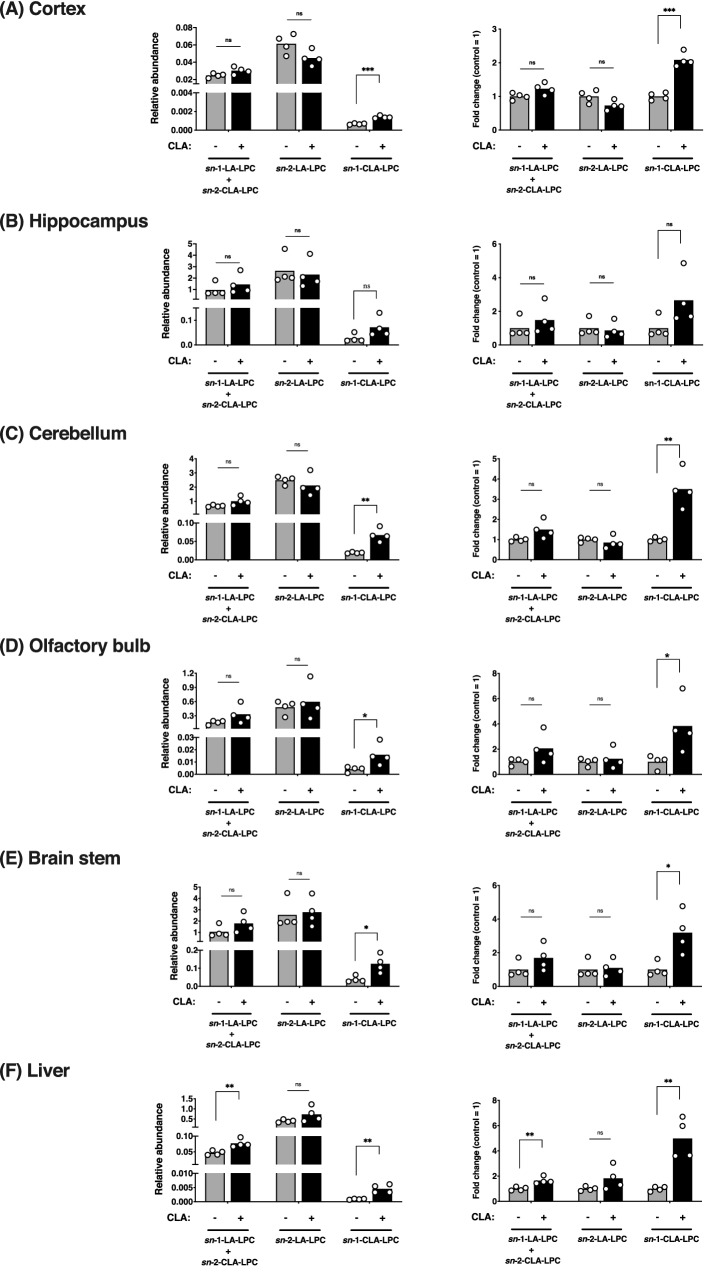
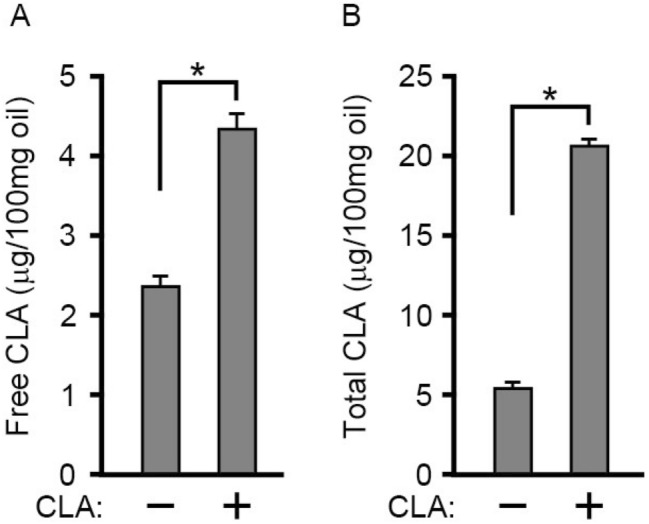
Conjugated linoleic acid (CLA) is an isomer of linoleic acid (LA). The predominant dietary CLA is cis-9, trans-11-CLA (c-9, t-11-CLA), which constitutes up to ~ 90% of total CLA and is thought to be responsible for the positive health benefits associated with CLA. However, the effects of c-9, t-11-CLA on Alzheimer's disease (AD) remain to be elucidated. In this study, we investigated the effect of dietary intake of c-9, t-11-CLA on the pathogenesis of an AD mouse model. We found that c-9, t-11-CLA diet-fed AD model mice significantly exhibited (1) a decrease in amyloid-β protein (Aβ) levels in the hippocampus, (2) an increase in the number of microglia, and (3) an increase in the number of astrocytes expressing the anti-inflammatory cytokines, interleukin-10 and 19 (IL-10, IL-19), with no change in the total number of astrocytes. In addition, liquid chromatography-tandem mass spectrometry (LC-MS/MS) and gas chromatographic analysis revealed that the levels of lysophosphatidylcholine (LPC) containing c-9, t-11-CLA (CLA-LPC) and free c-9, t-11-CLA were significantly increased in the brain of c-9, t-11-CLA diet-fed mice. Thus, dietary c-9, t-11-CLA entered the brain and appeared to exhibit beneficial effects on AD, including a decrease in Aβ levels and suppression of inflammation.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Masters CL, et al. Alzheimer’s disease. Nat. Rev. Dis. Prim. 2015;1:1–18. - PubMed
-
- Zou K, et al. Amyloid β-protein (Aβ)1–40 protects neurons from damage induced by Aβ1-42 in culture and in rat brain. J. Neurochem. 2003;87:609–619. - PubMed
-
- Reynolds CM, Roche HM. Conjugated linoleic acid and inflammatory cell signalling. Prostaglandins Leukot. Essent. Fat. Acids. 2010;82:199–204. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
