

Two-photon live imaging of single corneal stem cells reveals compartmentalized organization of the limbal niche
- PMID: 33984283
- PMCID: PMC8559309
- DOI: 10.1016/j.stem.2021.02.022
Two-photon live imaging of single corneal stem cells reveals compartmentalized organization of the limbal niche
Abstract
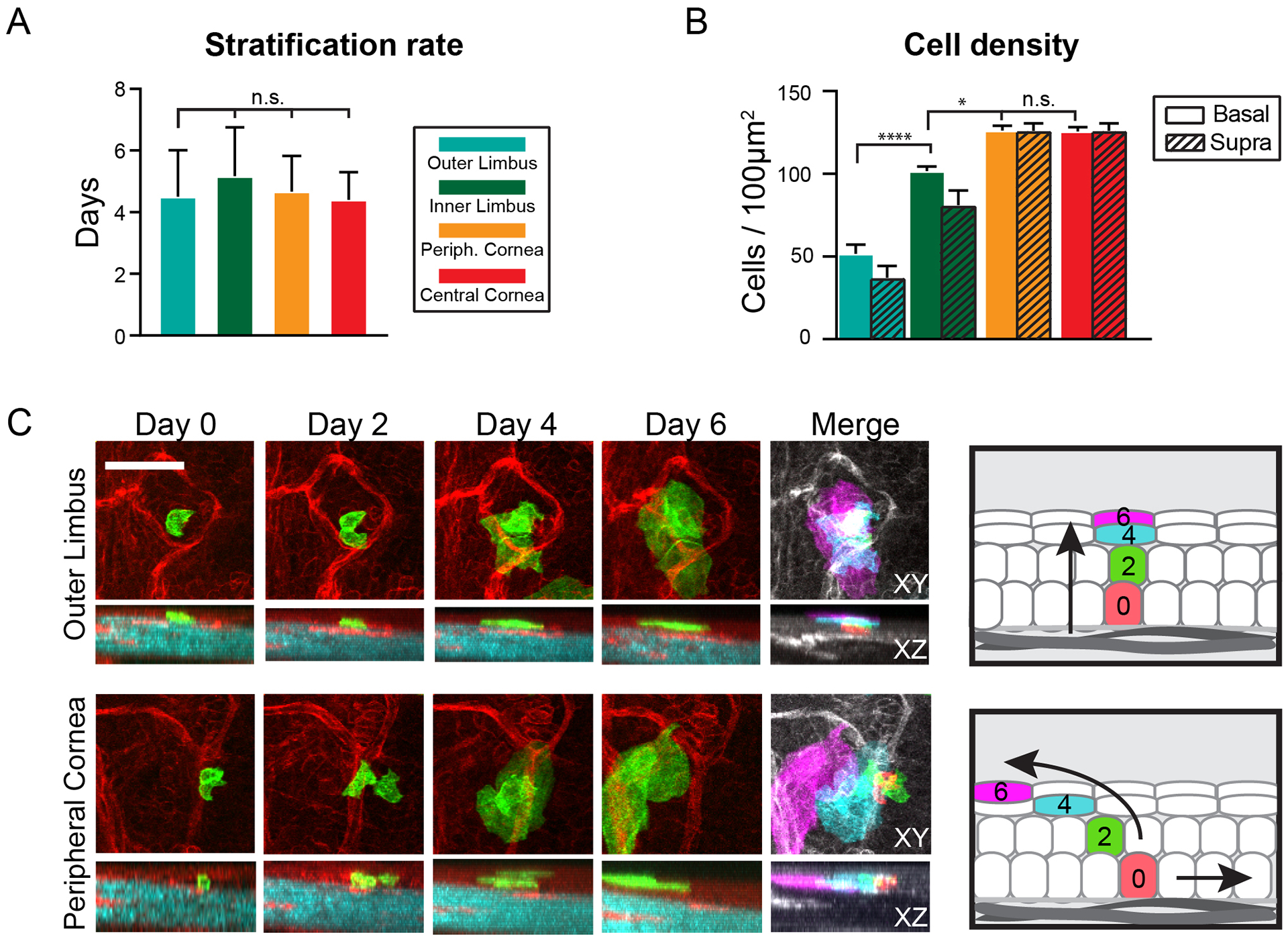
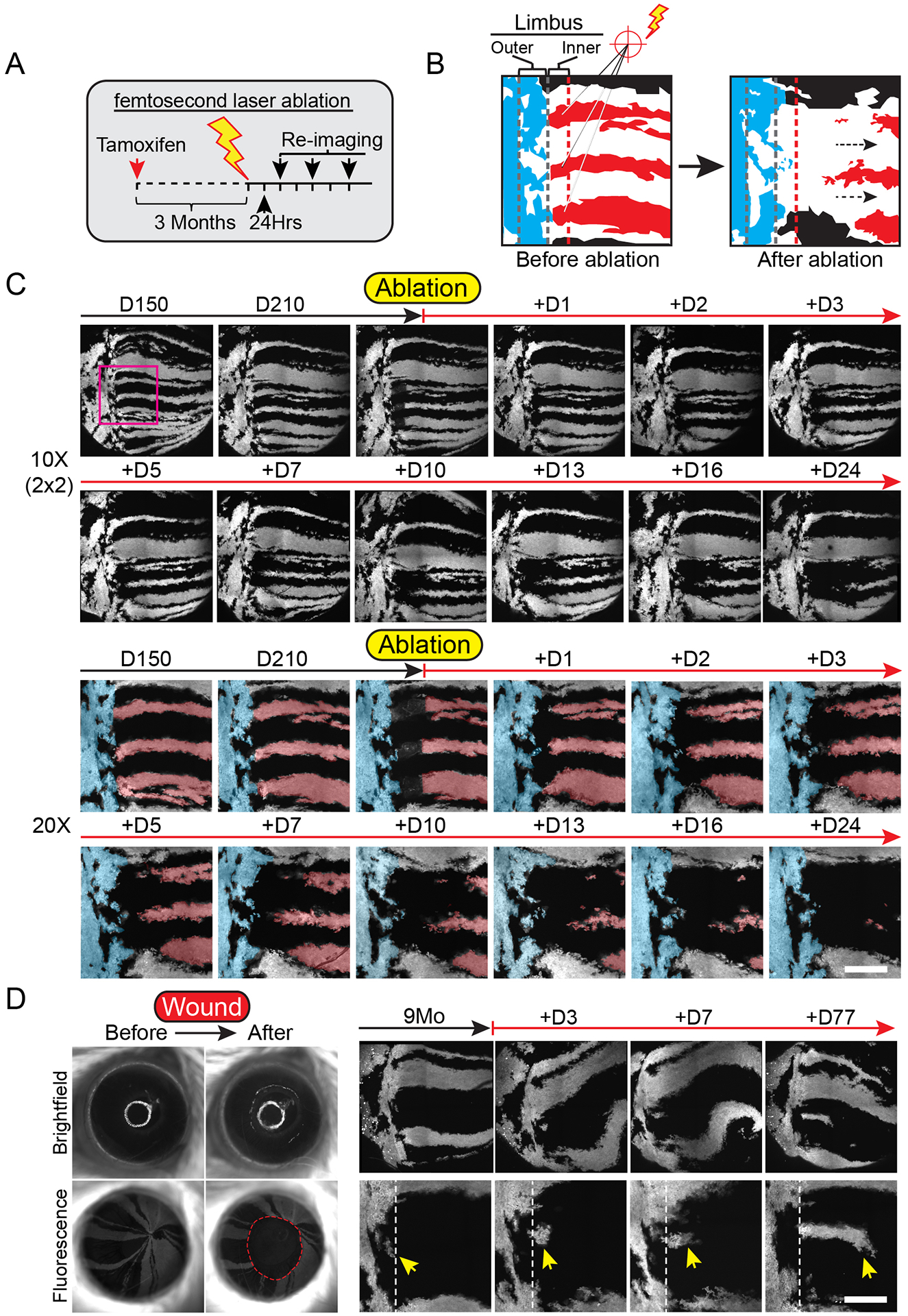
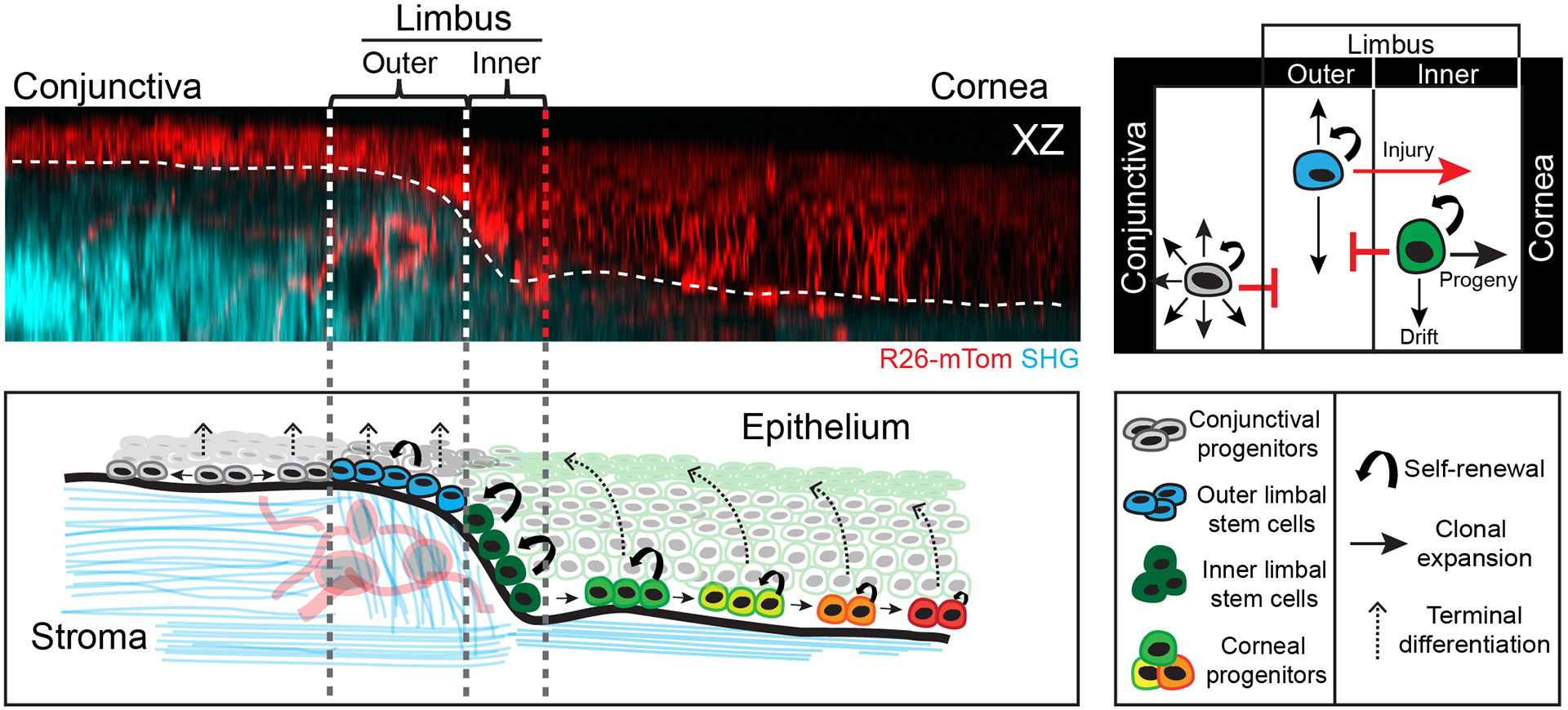
The functional heterogeneity of resident stem cells that support adult organs is incompletely understood. Here, we directly visualize the corneal limbus in the eyes of live mice and identify discrete stem cell niche compartments. By recording the life cycle of individual stem cells and their progeny, we directly analyze their fates and show that their location within the tissue can predict their differentiation status. Stem cells in the inner limbus undergo mostly symmetric divisions and are required to sustain the population of transient progenitors that support corneal homeostasis. Using in situ photolabeling, we captured their progeny exiting the niche before moving centripetally in unison. The long-implicated slow-cycling stem cells are functionally distinct and display local clonal dynamics during homeostasis but can contribute to corneal regeneration after injury. This study demonstrates how the compartmentalized organization of functionally diverse stem cell populations supports the maintenance and regeneration of an adult organ.
Keywords: cornea; epithelia; intravital imaging; limbus; niche; stem cells; two-photon microscopy.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
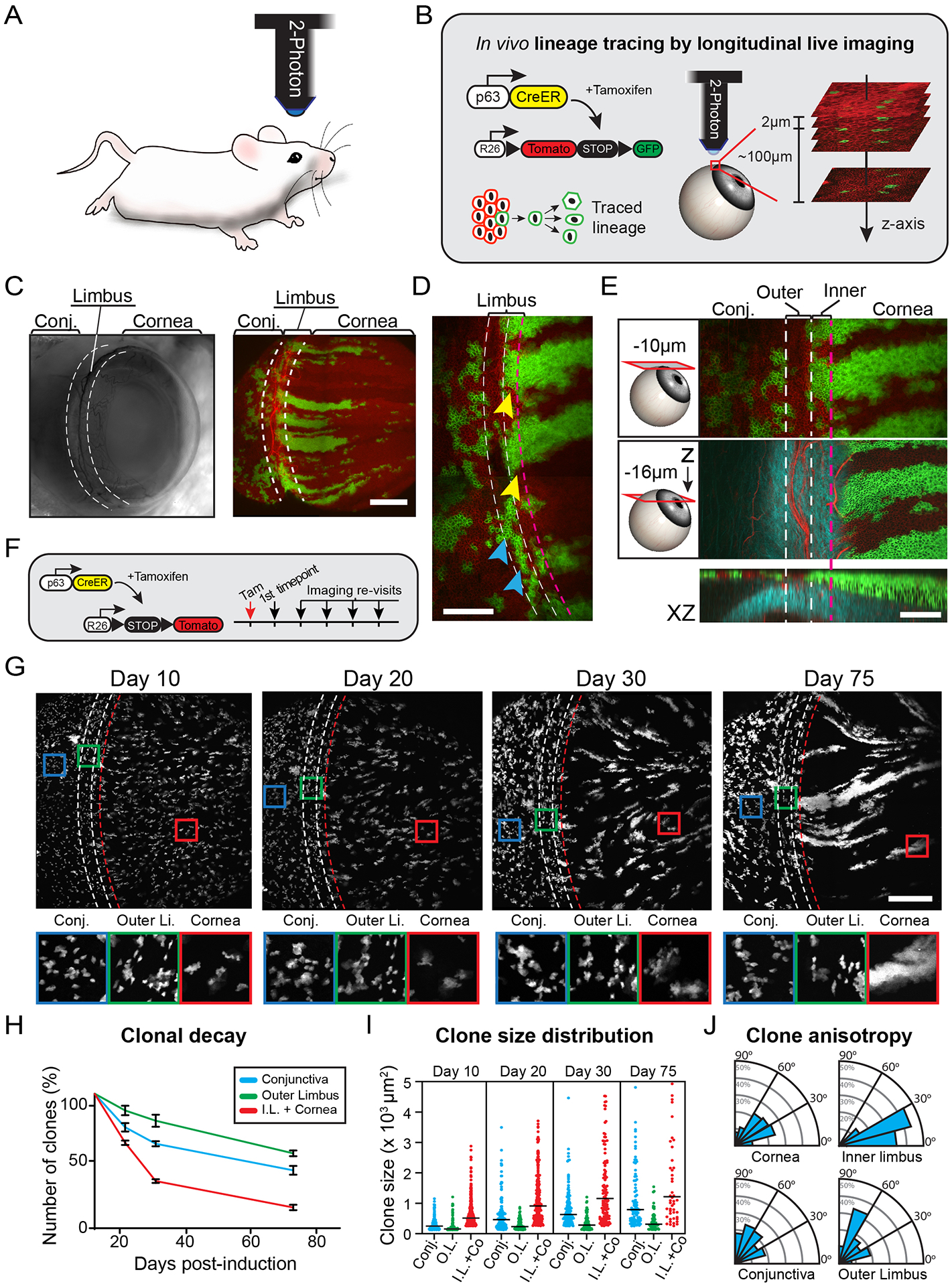
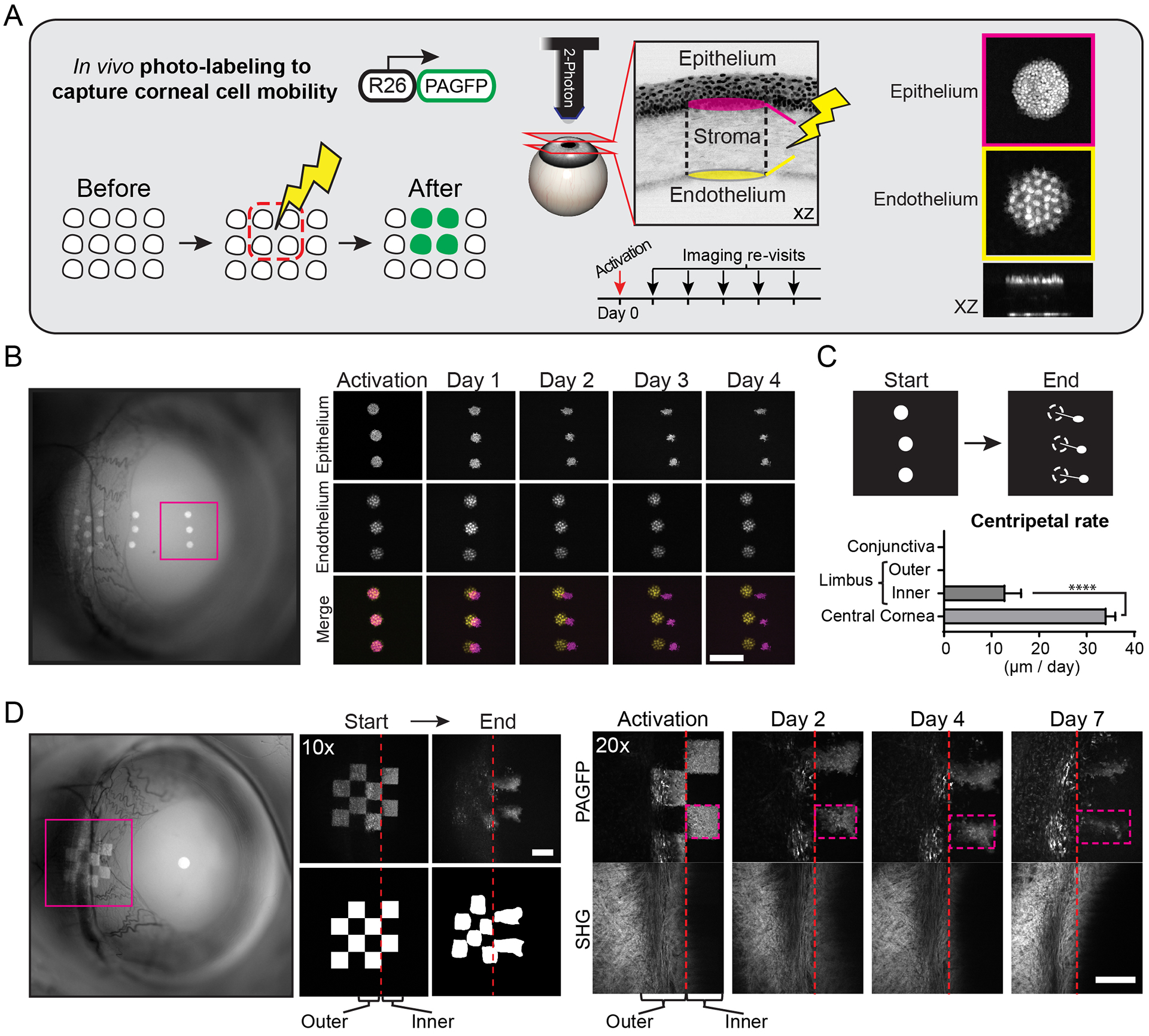
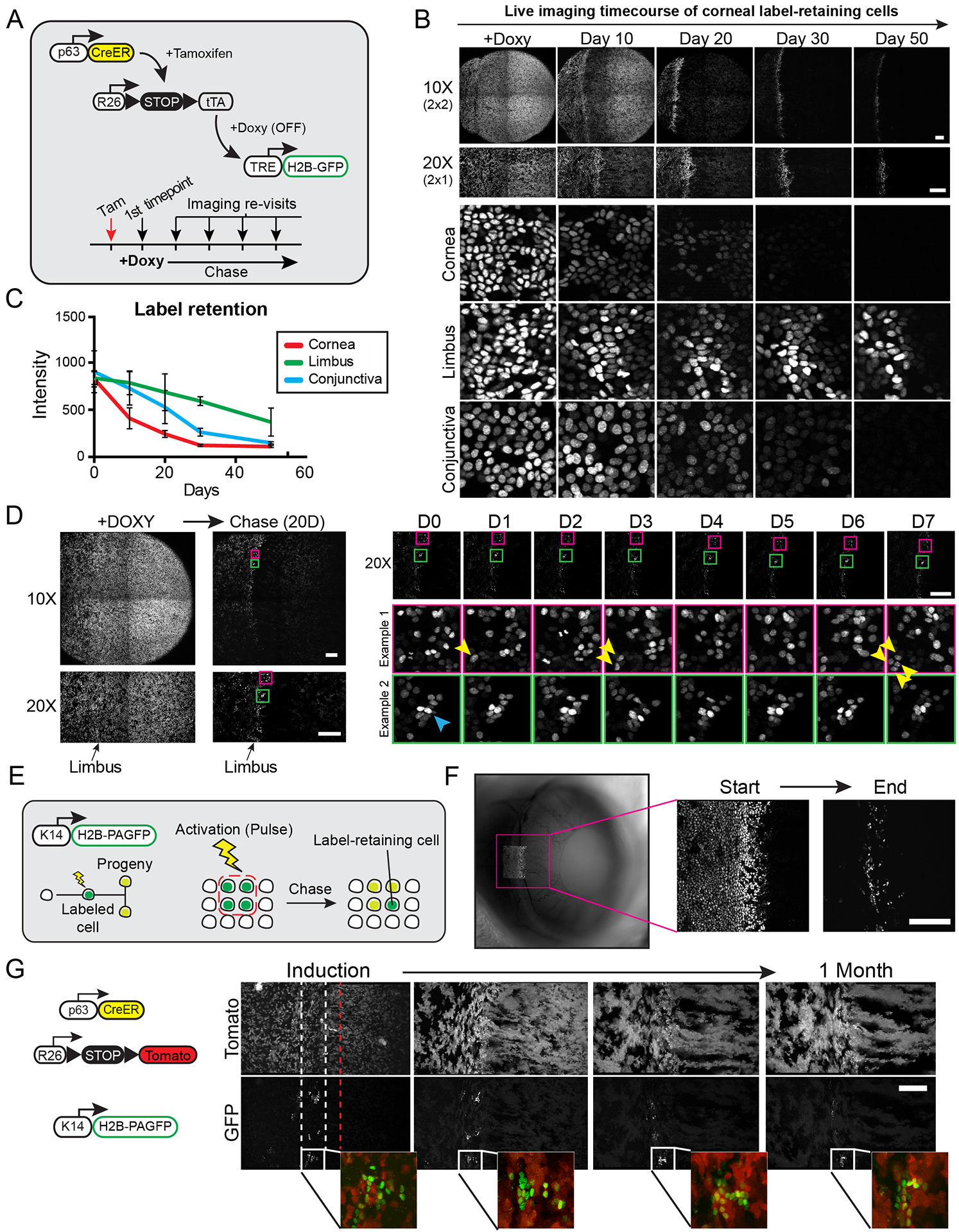
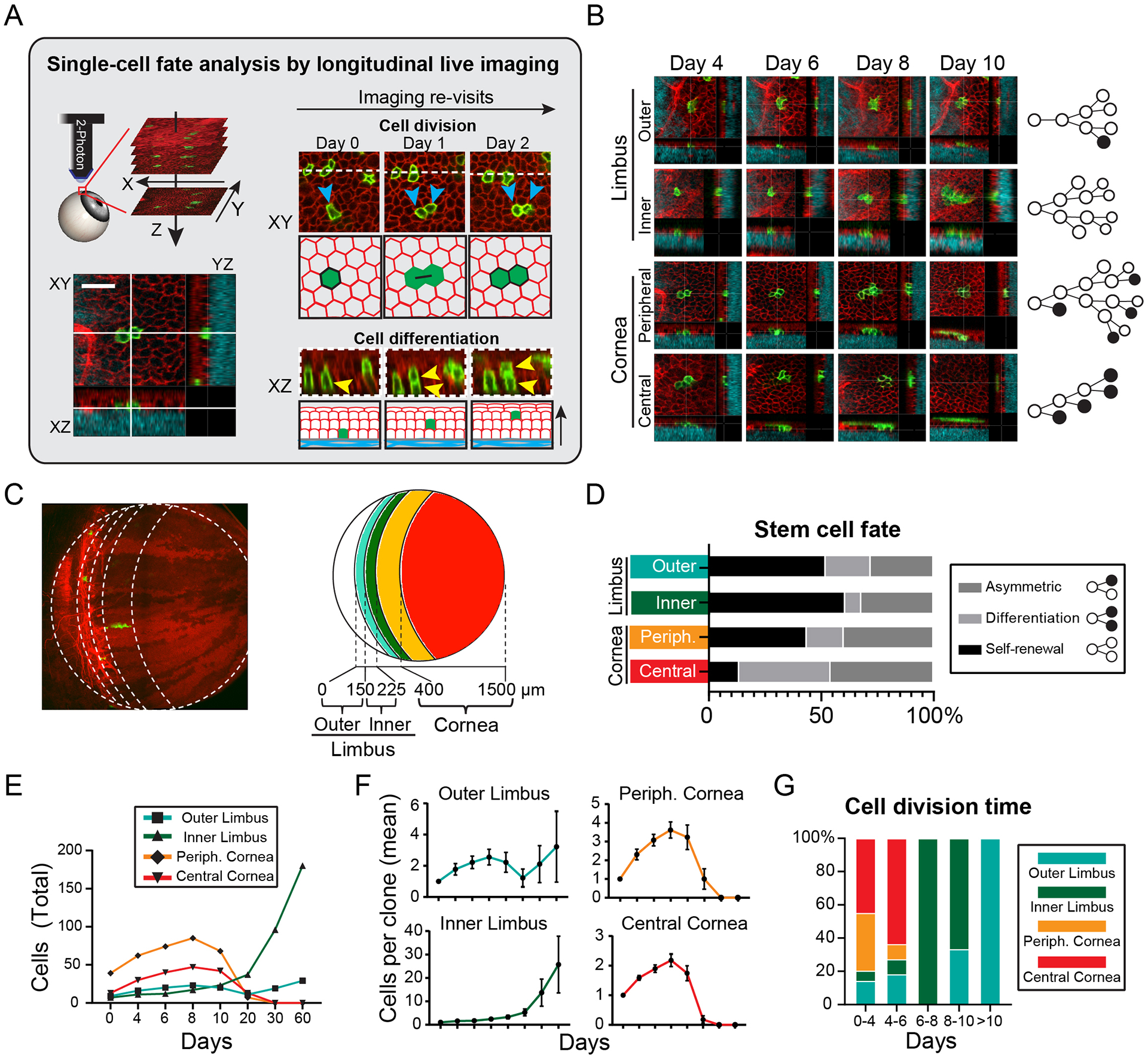
Comment in
-
"Eyeing" corneal stem cell identity, dynamics, and compartmentalization.Cell Stem Cell. 2021 Jul 1;28(7):1181-1183. doi: 10.1016/j.stem.2021.06.001. Cell Stem Cell. 2021. PMID: 34214435
References
-
- Amitai-Lange A, Altshuler A, et al., 2015. Lineage tracing of stem and progenitor cells of the murine corneal epithelium. Stem cells (Dayton, Ohio), 33(1), pp. 230–239. - PubMed
-
- Busch K et al., 2015. Fundamental properties of unperturbed haematopoiesis from stem cells in vivo. Nature, 518(7540), pp. 542–546. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
