

Plant-derived exosomal microRNAs inhibit lung inflammation induced by exosomes SARS-CoV-2 Nsp12
- PMID: 33984520
- PMCID: PMC8110335
- DOI: 10.1016/j.ymthe.2021.05.005
Plant-derived exosomal microRNAs inhibit lung inflammation induced by exosomes SARS-CoV-2 Nsp12
Abstract
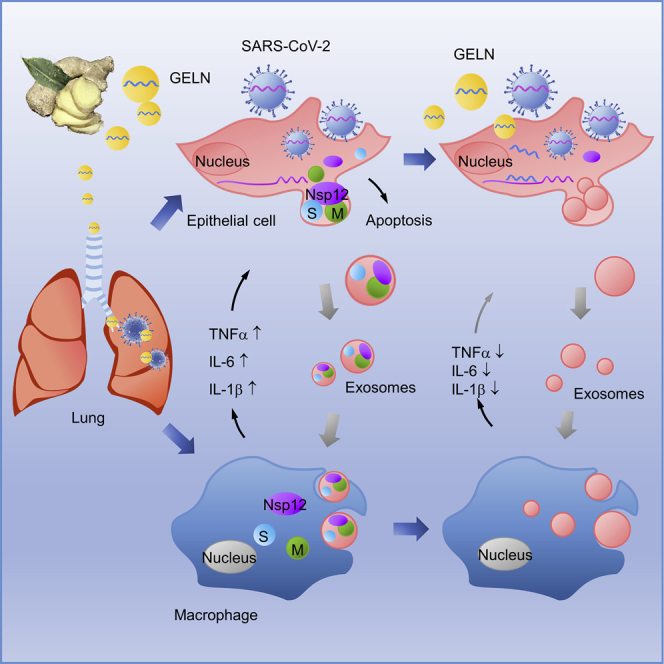
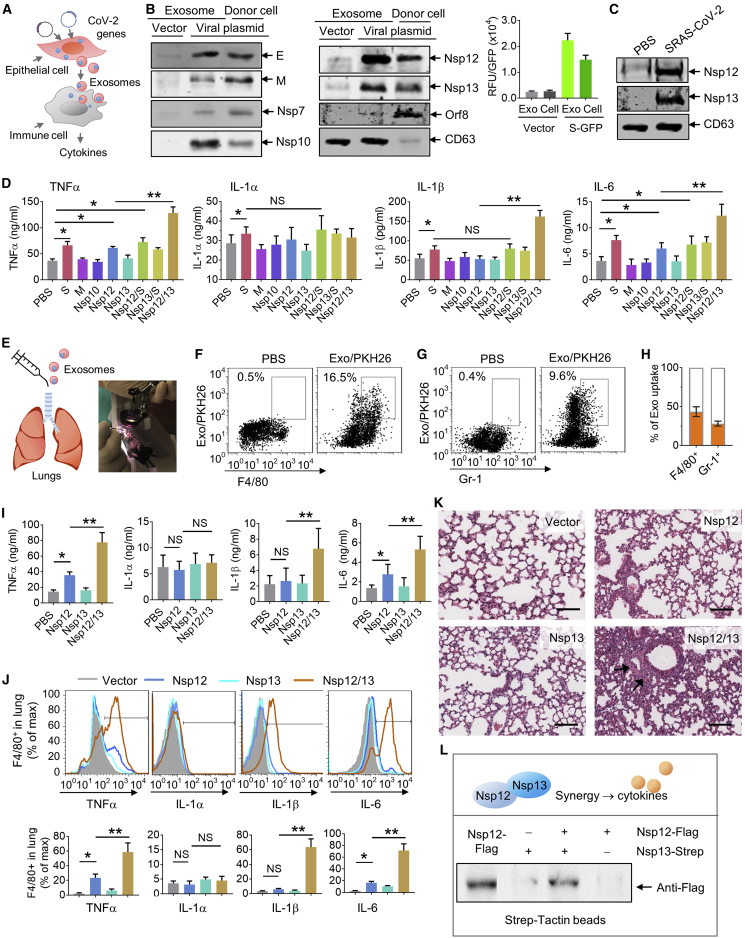
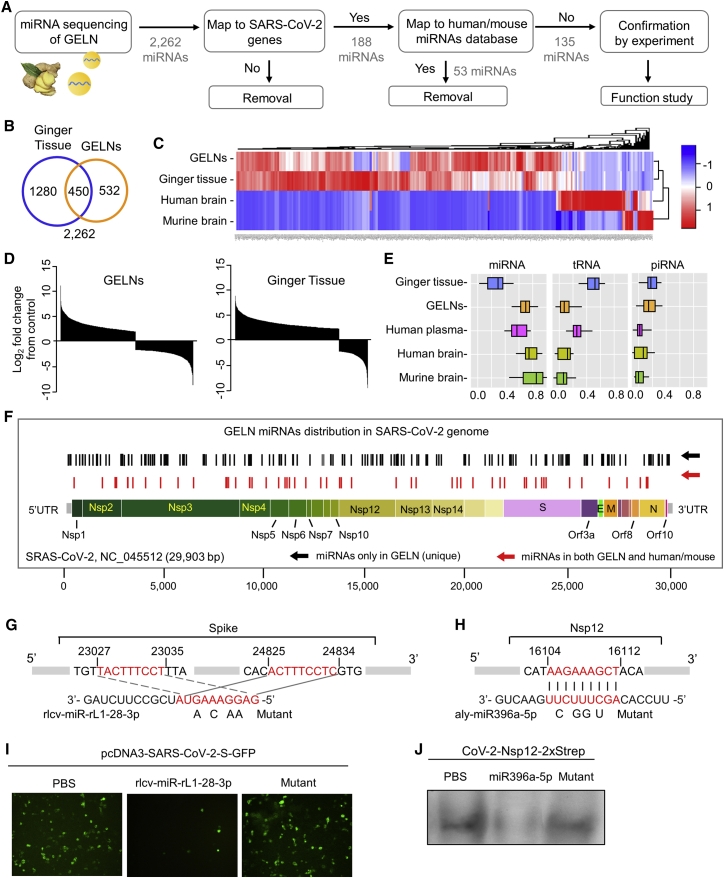
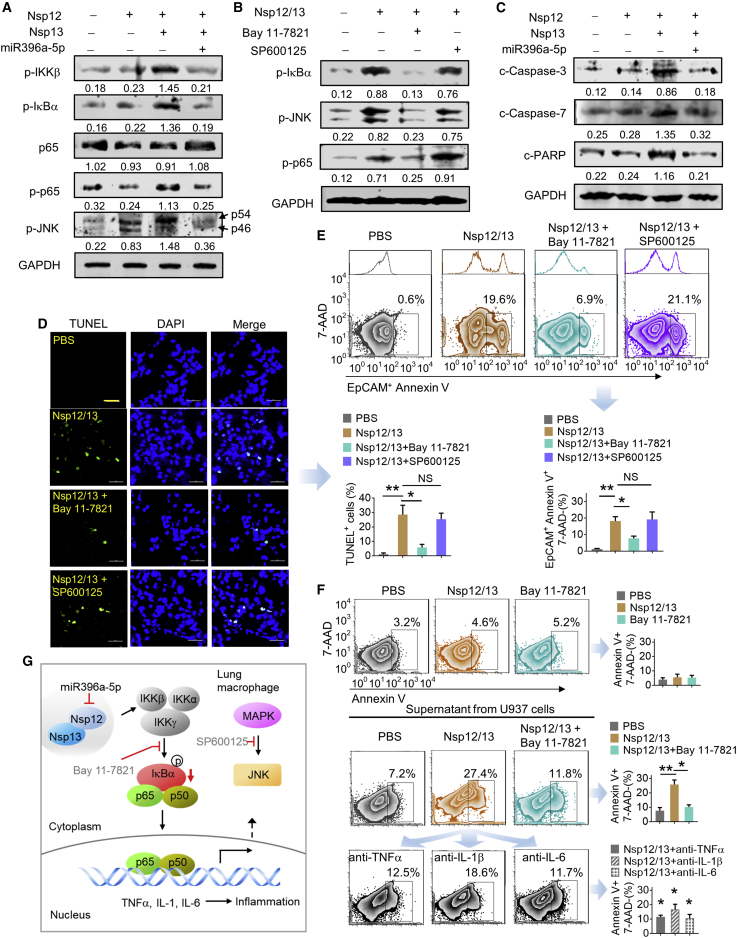
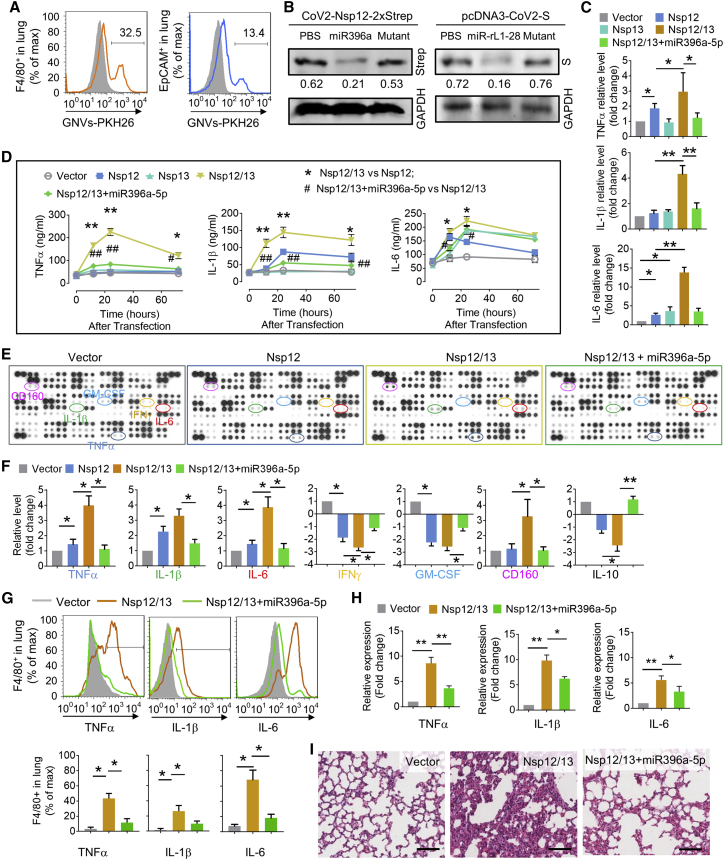
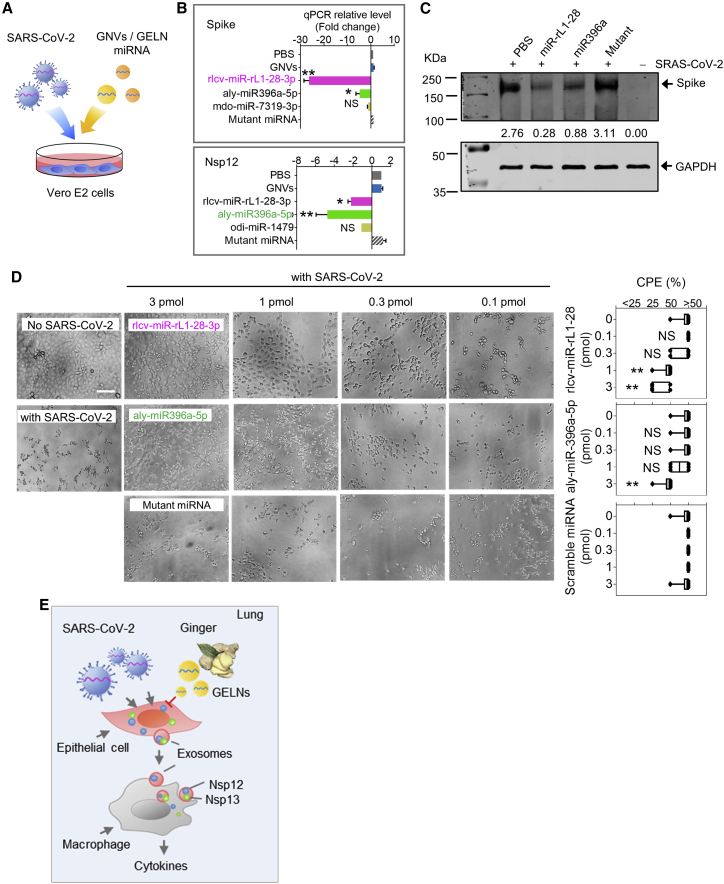
Lung inflammation is a hallmark of coronavirus disease 2019 (COVID-19). In this study, we show that mice develop inflamed lung tissue after being administered exosomes released from the lung epithelial cells exposed to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) Nsp12 and Nsp13 (exosomesNsp12Nsp13). Mechanistically, we show that exosomesNsp12Nsp13 are taken up by lung macrophages, leading to activation of nuclear factor κB (NF-κB) and the subsequent induction of an array of inflammatory cytokines. Induction of tumor necrosis factor (TNF)-α, interleukin (IL)-6, and IL-1β from exosomesNsp12Nsp13-activated lung macrophages contributes to inducing apoptosis in lung epithelial cells. Induction of exosomesNsp12Nsp13-mediated lung inflammation was abolished with ginger exosome-like nanoparticle (GELN) microRNA (miRNA aly-miR396a-5p. The role of GELNs in inhibition of the SARS-CoV-2-induced cytopathic effect (CPE) was further demonstrated via GELN aly-miR396a-5p- and rlcv-miR-rL1-28-3p-mediated inhibition of expression of Nsp12 and spike genes, respectively. Taken together, our results reveal exosomesNsp12Nsp13 as potentially important contributors to the development of lung inflammation, and GELNs are a potential therapeutic agent to treat COVID-19.
Keywords: ACE2; IL-1β; IL-6; NF-κB; Nsp12; SARS-CoV-2; TNF-α; exosomes; ginger exosome-like nanoparticle; lung epithelial cells; lung inflammation; macrophages; microRNA; spike.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
