

Engineering complex communities by directed evolution
- PMID: 33986540
- PMCID: PMC8263491
- DOI: 10.1038/s41559-021-01457-5
Engineering complex communities by directed evolution
Abstract
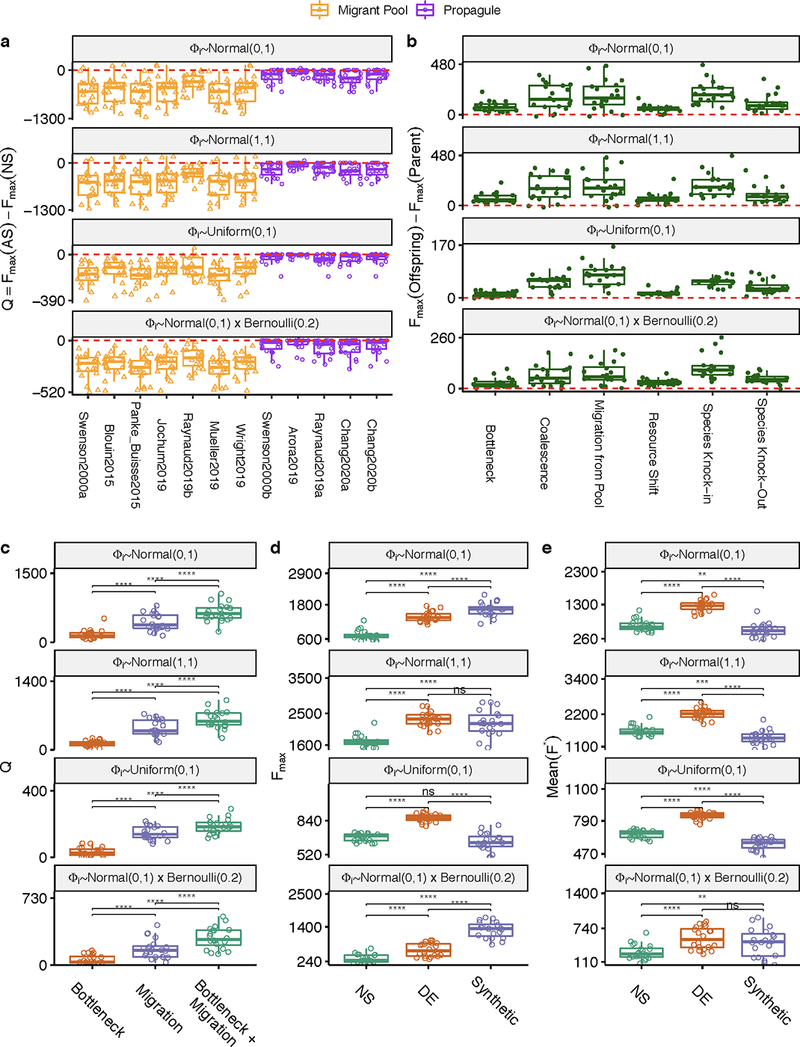
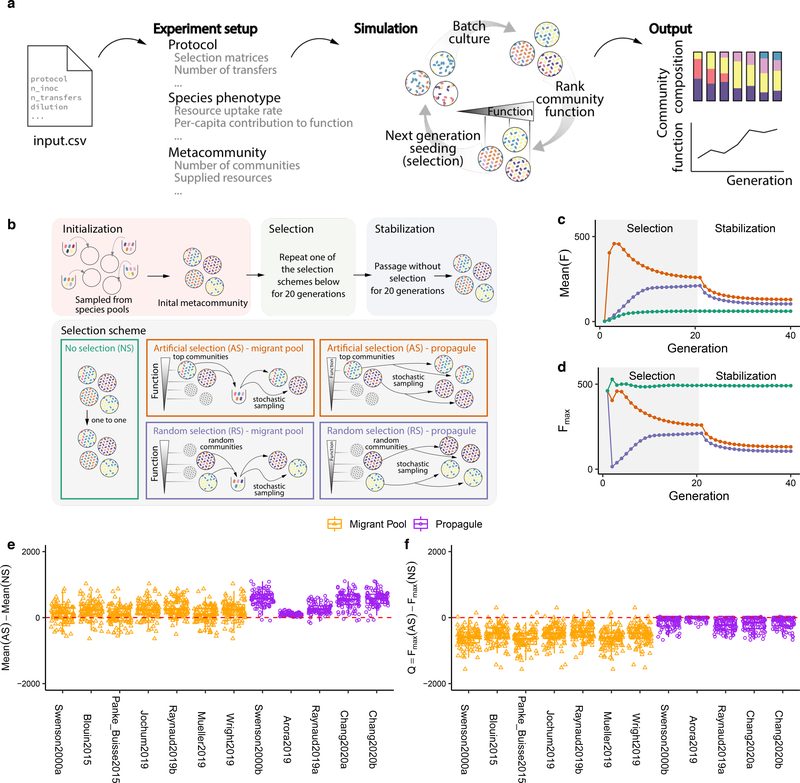
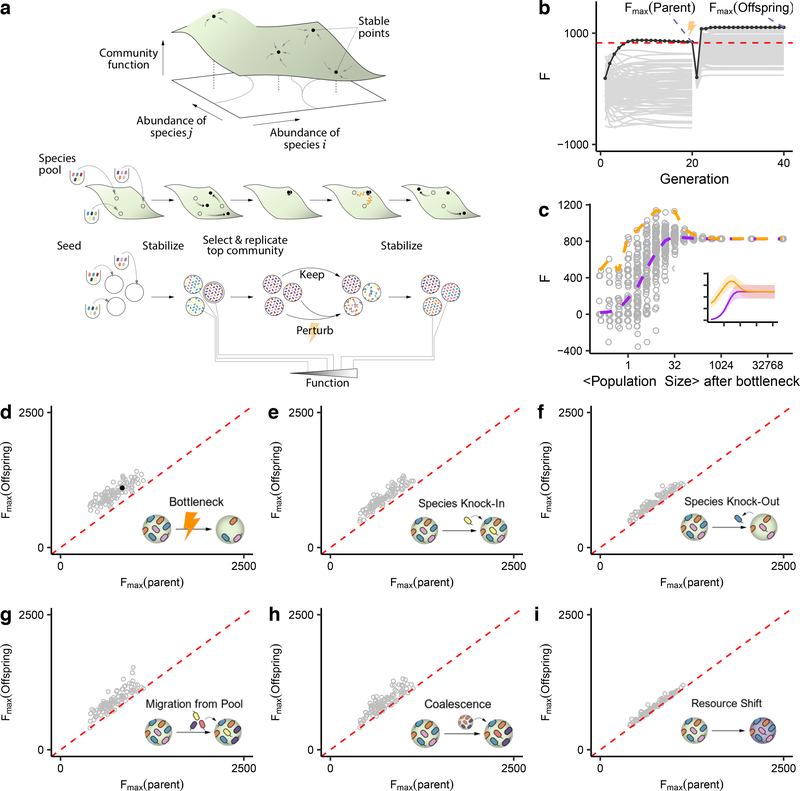
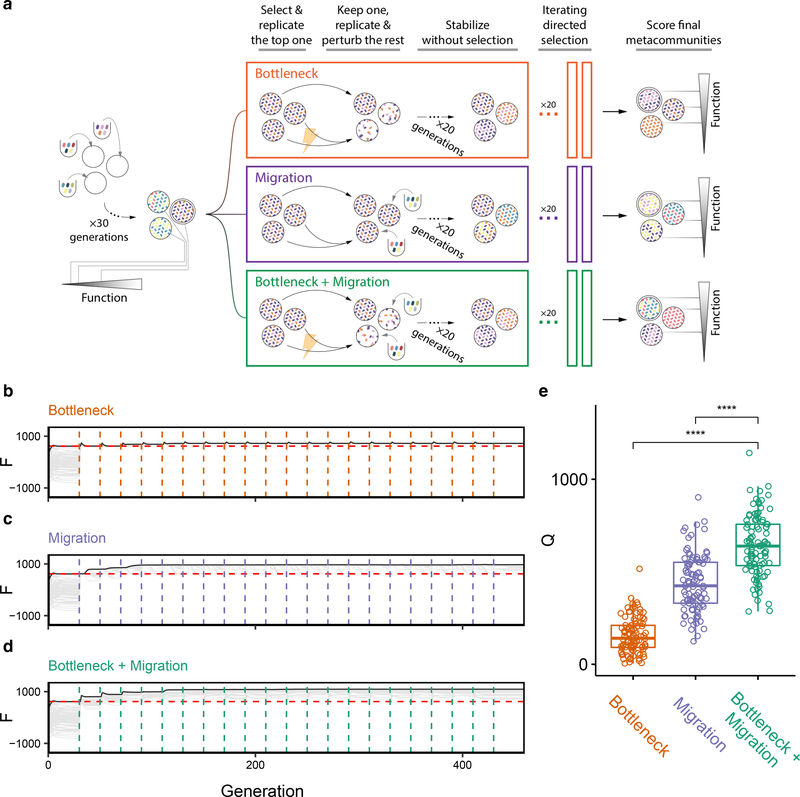
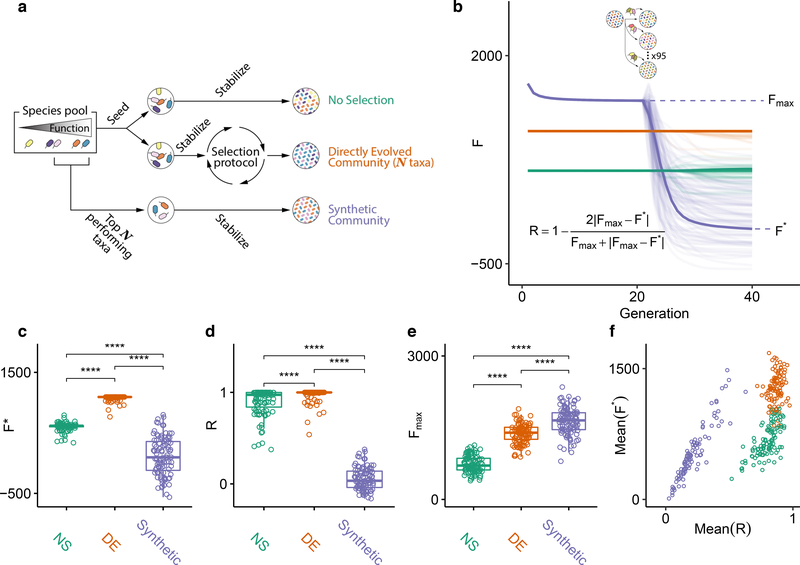
Directed evolution has been used for decades to engineer biological systems at or below the organismal level. Above the organismal level, a small number of studies have attempted to artificially select microbial ecosystems, with uneven and generally modest success. Our theoretical understanding of artificial ecosystem selection is limited, particularly for large assemblages of asexual organisms, and we know little about designing efficient methods to direct their evolution. Here, we have developed a flexible modelling framework that allows us to systematically probe any arbitrary selection strategy on any arbitrary set of communities and selected functions. By artificially selecting hundreds of in silico microbial metacommunities under identical conditions, we first show that the main breeding methods used to date, which do not necessarily let communities reach their ecological equilibrium, are outperformed by a simple screen of sufficiently mature communities. We then identify a range of alternative directed evolution strategies that, particularly when applied in combination, are well suited for the top-down engineering of large, diverse and stable microbial consortia. Our results emphasize that directed evolution allows an ecological structure-function landscape to be navigated in search of dynamically stable and ecologically resilient communities with desired quantitative attributes.
Conflict of interest statement
Figures
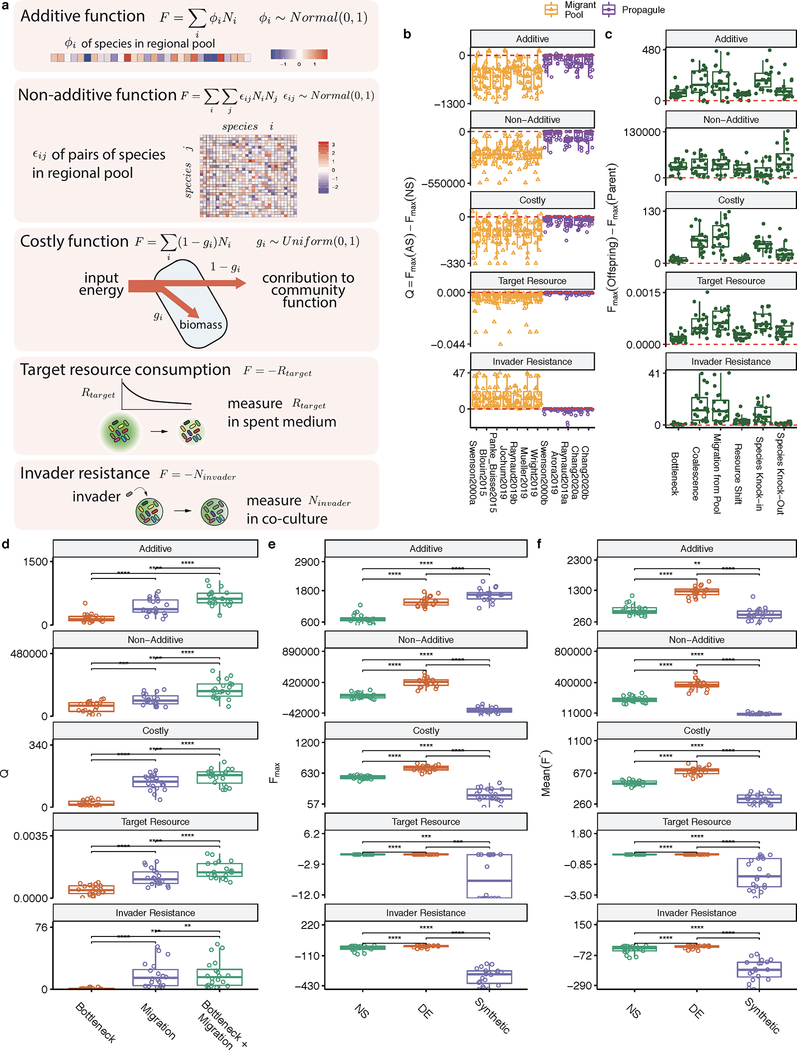
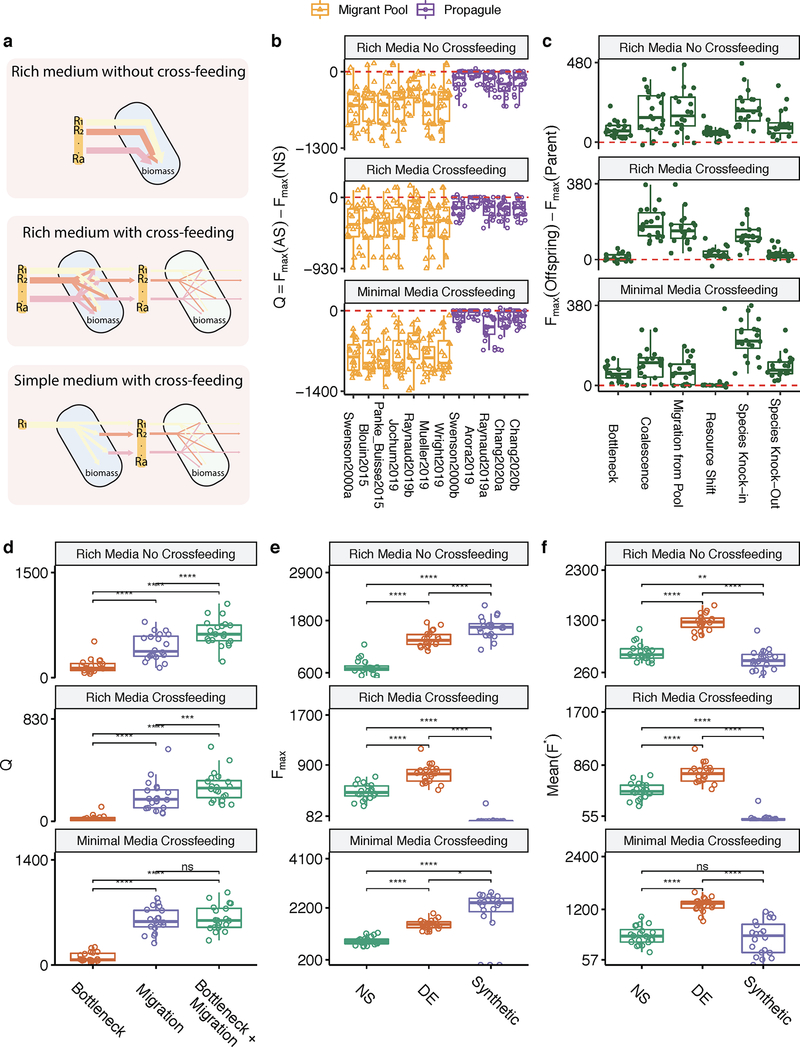
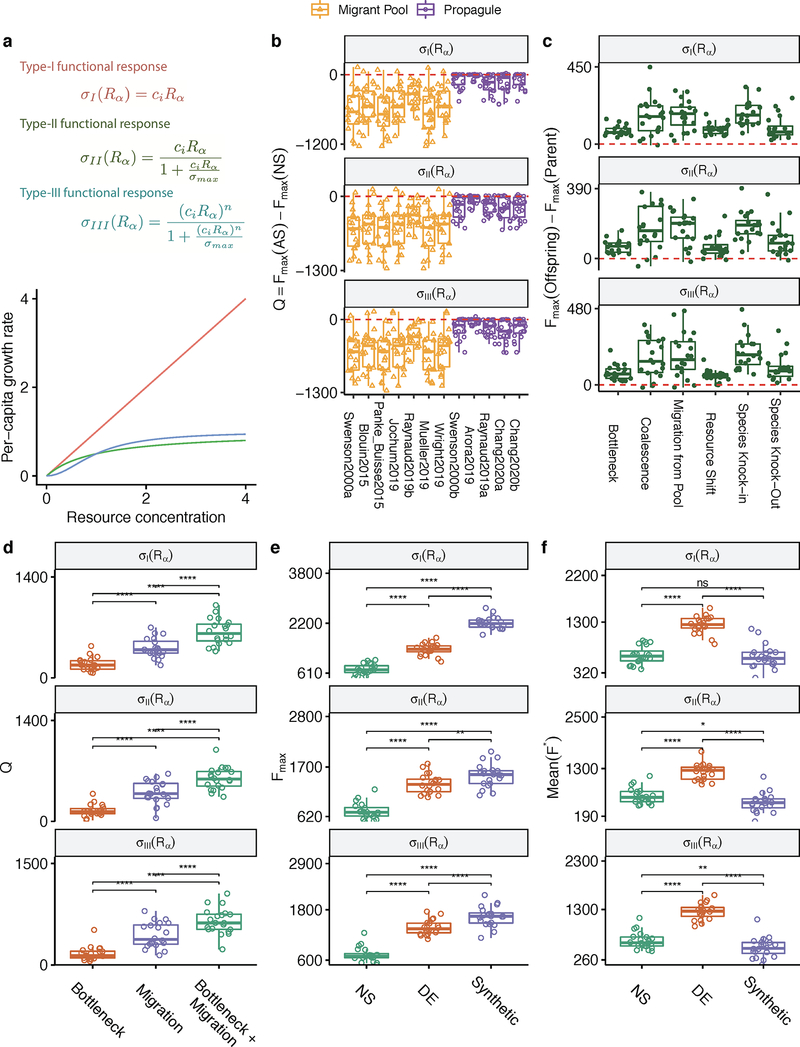
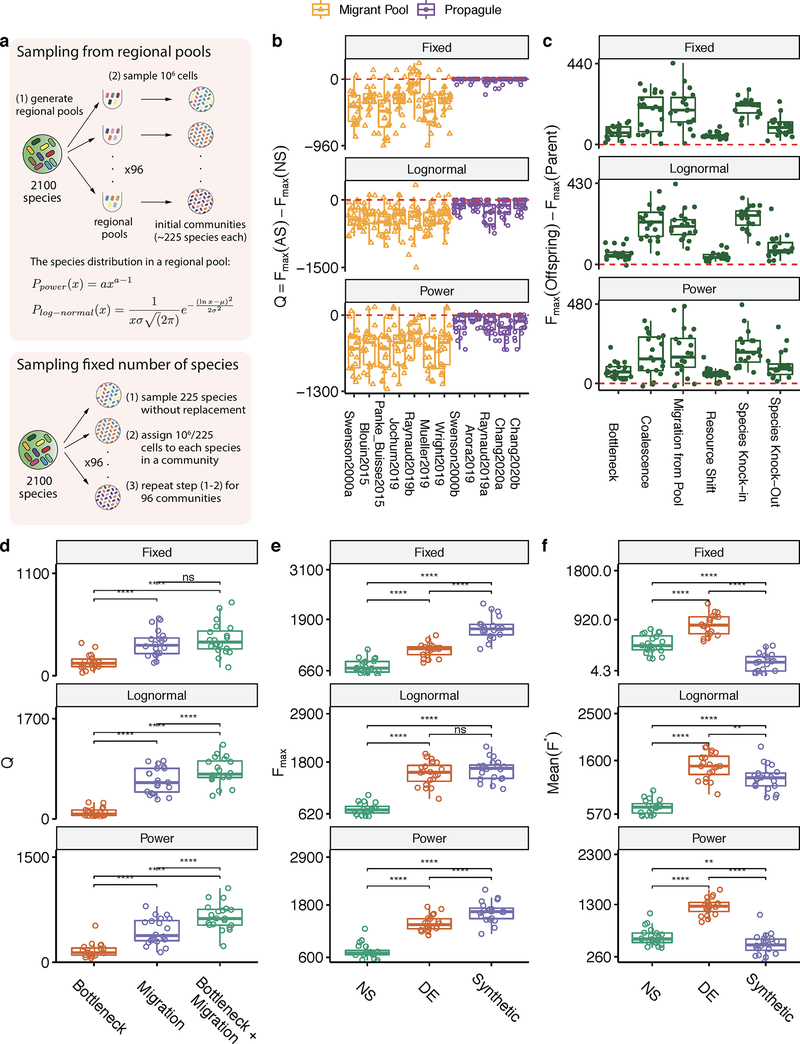
Comment in
-
Ecological recipes for selecting community function.Nat Ecol Evol. 2021 Jul;5(7):894-895. doi: 10.1038/s41559-021-01467-3. Nat Ecol Evol. 2021. PMID: 33986539 No abstract available.
References
-
- Mueller UG & Sachs JL Engineering Microbiomes to Improve Plant and Animal Health. Trends Microbiol 23, 606–617 (2015). - PubMed
-
- Gilbert ES, Walker AW & Keasling JD A constructed microbial consortium for biodegradation of the organophosphorus insecticide parathion. Appl. Microbiol. Biotechnol 61, 77–81 (2003). - PubMed
-
- Yoshida S, Ogawa N, Fujii T & Tsushima S Enhanced biofilm formation and 3-chlorobenzoate degrading activity by the bacterial consortium of Burkholderia sp. NK8 and Pseudomonas aeruginosa PAO1. J. Appl. Microbiol 106, 790–800 (2009). - PubMed
