

Adult neurogenic process in the subventricular zone-olfactory bulb system is regulated by Tau protein under prolonged stress
- PMID: 33988263
- PMCID: PMC8249793
- DOI: 10.1111/cpr.13027
Adult neurogenic process in the subventricular zone-olfactory bulb system is regulated by Tau protein under prolonged stress
Abstract
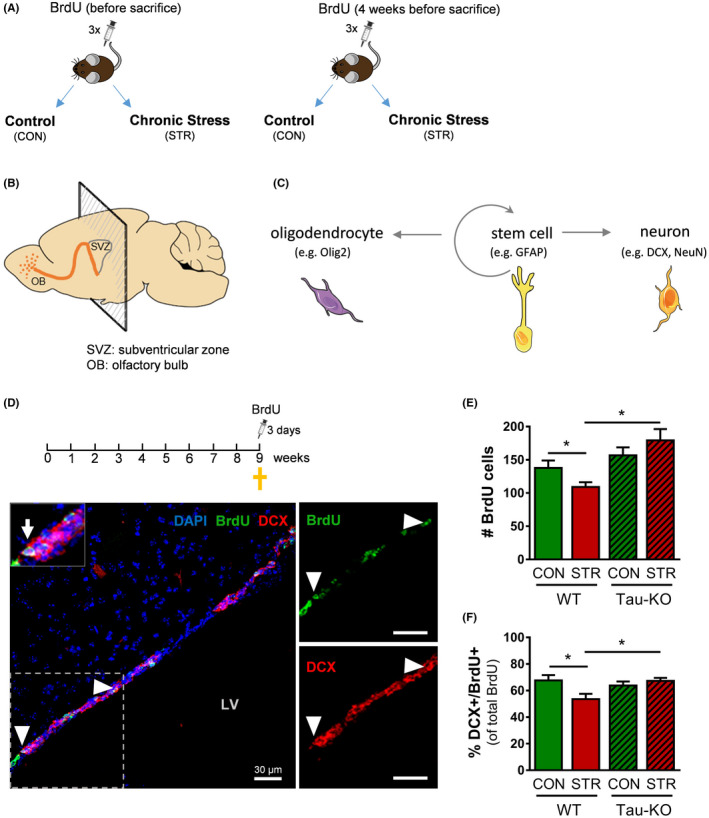
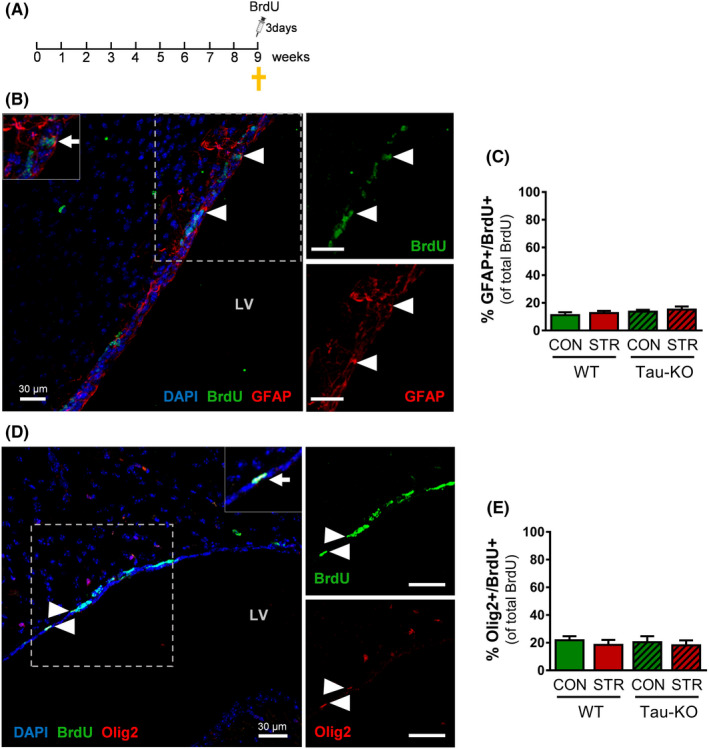
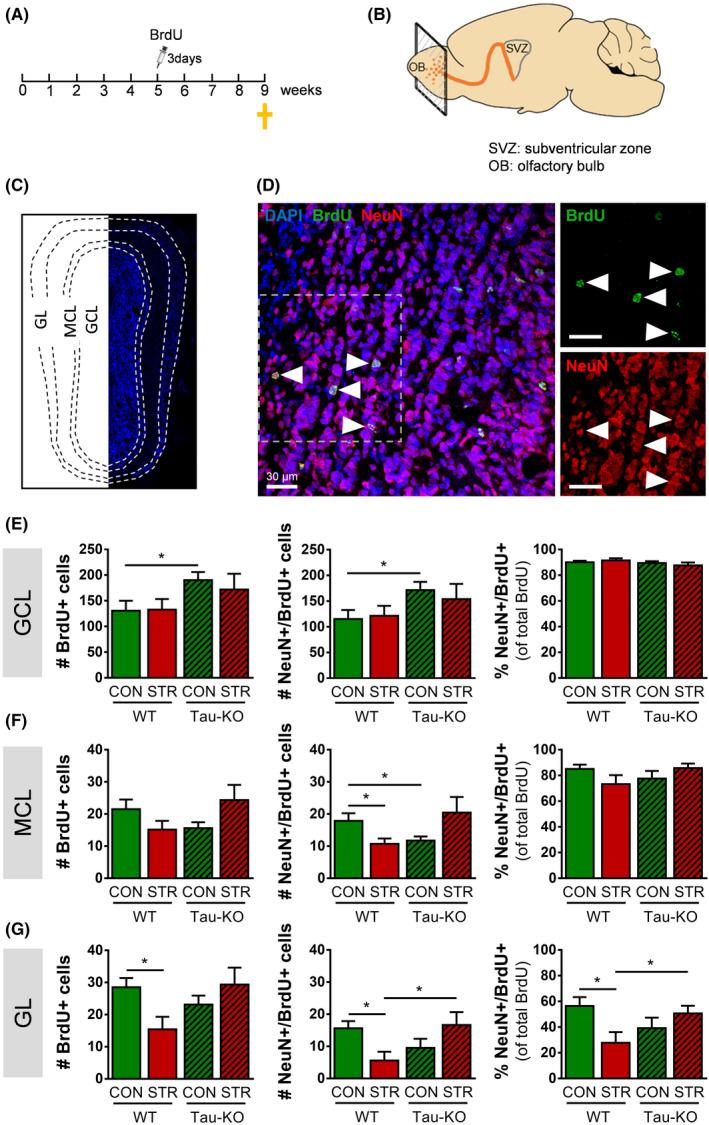
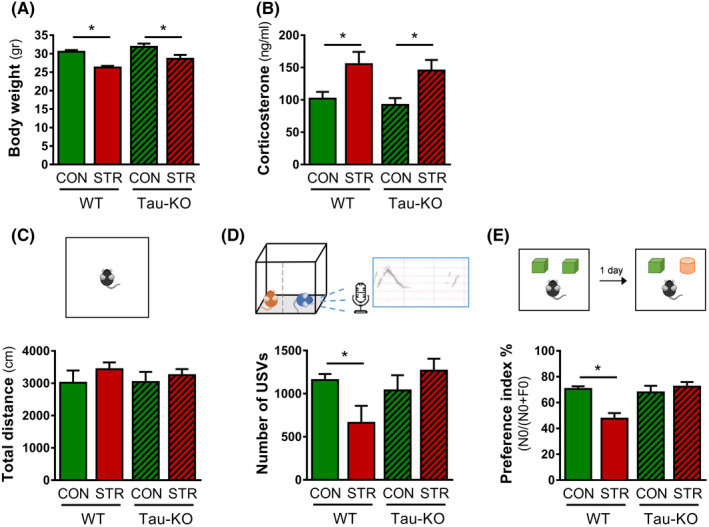
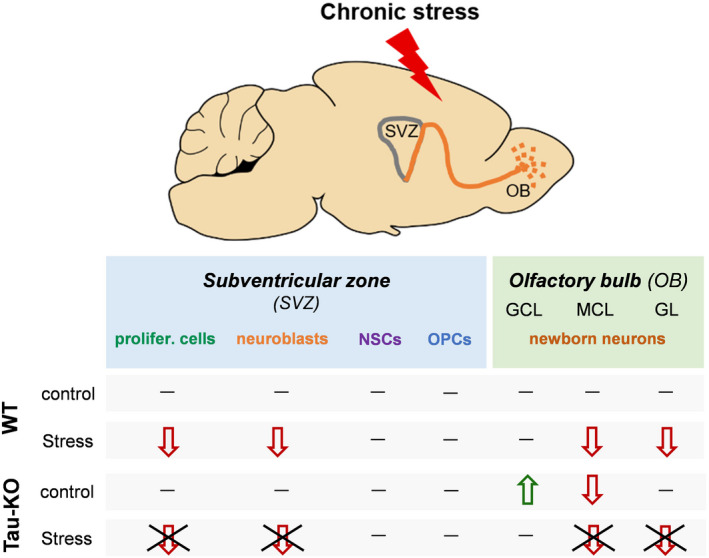
Objectives: The area of the subventricular zone (SVZ) in the adult brain exhibits the highest number of proliferative cells, which, together with the olfactory bulb (OB), maintains constant brain plasticity through the generation, migration and integration of newly born neurons. Despite Tau and its malfunction is increasingly related to deficits of adult hippocampal neurogenesis and brain plasticity under pathological conditions [e.g. in Alzheimer's disease (AD)], it remains unknown whether Tau plays a role in the neurogenic process of the SVZ and OB system under conditions of chronic stress, a well-known sculptor of brain and risk factor for AD.
Materials and methods: Different types of newly born cells in SVZ and OB were analysed in animals that lack Tau gene (Tau-KO) and their wild-type littermates (WT) under control or chronic stress conditions.
Results: We demonstrate that chronic stress reduced the number of proliferating cells and neuroblasts in the SVZ leading to decreased number of newborn neurons in the OB of adult WT, but not Tau-KO, mice. Interestingly, while stress-evoked changes were not detected in OB granular cell layer, Tau-KO exhibited increased number of mature neurons in this layer indicating altered neuronal migration due to Tau loss.
Conclusions: Our findings suggest the critical involvement of Tau in the neurogenesis suppression of SVZ and OB neurogenic niche under stressful conditions highlighting the role of Tau protein as an essential regulator of stress-driven plasticity deficits.
Keywords: Tau protein; chronic stress; neurogenesis; olfactory bulb; oligodendrogenesis; subventricular zone.
© 2021 The Authors. Cell Proliferation published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Running-Activated Neural Stem Cells Enhance Subventricular Neurogenesis and Improve Olfactory Behavior in p21 Knockout Mice.Mol Neurobiol. 2019 Nov;56(11):7534-7556. doi: 10.1007/s12035-019-1590-6. Epub 2019 May 6. Mol Neurobiol. 2019. PMID: 31062248
-
Drebrin E regulates neuroblast proliferation and chain migration in the adult brain.Eur J Neurosci. 2017 Sep;46(6):2214-2228. doi: 10.1111/ejn.13668. Epub 2017 Sep 7. Eur J Neurosci. 2017. PMID: 28833685
-
Calorie restriction activates new adult born olfactory-bulb neurones in a ghrelin-dependent manner but acyl-ghrelin does not enhance subventricular zone neurogenesis.J Neuroendocrinol. 2019 Jul;31(7):e12755. doi: 10.1111/jne.12755. Epub 2019 Jun 24. J Neuroendocrinol. 2019. PMID: 31179562
-
Olfactory bulb neurogenesis depending on signaling in the subventricular zone.Cereb Cortex. 2023 Nov 4;33(22):11102-11111. doi: 10.1093/cercor/bhad349. Cereb Cortex. 2023. PMID: 37746807 Review.
-
Dynamic changes in the transcriptional profile of subventricular zone-derived postnatally born neuroblasts.Mech Dev. 2013 Jun-Aug;130(6-8):424-32. doi: 10.1016/j.mod.2012.11.003. Epub 2012 Dec 5. Mech Dev. 2013. PMID: 23220001 Review.
Cited by
-
Neurogenesis in aging and age-related neurodegenerative diseases.Ageing Res Rev. 2022 Jun;78:101636. doi: 10.1016/j.arr.2022.101636. Epub 2022 Apr 29. Ageing Res Rev. 2022. PMID: 35490966 Free PMC article. Review.
-
Melatonin Supplementation in Alzheimer's disease: The Potential Role in Neurogenesis.Mol Neurobiol. 2025 May 29. doi: 10.1007/s12035-025-05095-x. Online ahead of print. Mol Neurobiol. 2025. PMID: 40439856 Review.
-
The Mitochondrial Antioxidant Sirtuin3 Cooperates with Lipid Metabolism to Safeguard Neurogenesis in Aging and Depression.Cells. 2021 Dec 29;11(1):90. doi: 10.3390/cells11010090. Cells. 2021. PMID: 35011652 Free PMC article.
-
Sustaining Brain Youth by Neural Stem Cells: Physiological and Therapeutic Perspectives.Mol Neurobiol. 2025 Jul;62(7):8222-8247. doi: 10.1007/s12035-025-04774-z. Epub 2025 Feb 22. Mol Neurobiol. 2025. PMID: 39985708 Free PMC article. Review.
-
Relationship between adult subventricular neurogenesis and Alzheimer's disease: Pathologic roles and therapeutic implications.Front Aging Neurosci. 2022 Sep 14;14:1002281. doi: 10.3389/fnagi.2022.1002281. eCollection 2022. Front Aging Neurosci. 2022. PMID: 36185481 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
- FCOMP-01-0124-FEDER-037298/European Regional Development Fund
- POCI-01-0145-FEDER-007038/European Regional Development Fund
- CEECIND/02231/2018/Fundação para a Ciência e a Tecnologia
- IF/01079/2014/Fundação para a Ciência e a Tecnologia
- NORTE-01-0145-FEDER-000013/Fundação para a Ciência e a Tecnologia
- NORTE-01-0145-FEDER-000023/Fundação para a Ciência e a Tecnologia
- PEst-C/SAU/LA0026/2013/Fundação para a Ciência e a Tecnologia
- UIDB/50026/2020/Fundação para a Ciência e a Tecnologia
- UIDP/50026/2020/Fundação para a Ciência e a Tecnologia
- PPBI-POCI-01-0145-FEDER-022122/Portuguese Platform of Bioimaging
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
