

Inbreeding in a dioecious plant has sex- and population origin-specific effects on its interactions with pollinators
- PMID: 33988502
- PMCID: PMC8159375
- DOI: 10.7554/eLife.65610
Inbreeding in a dioecious plant has sex- and population origin-specific effects on its interactions with pollinators
Abstract
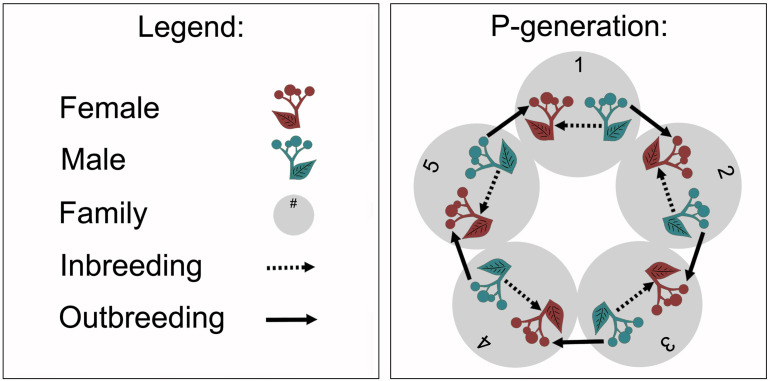
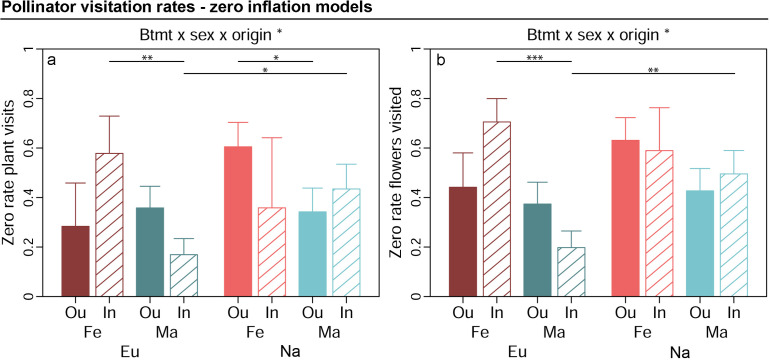
We study the effects of inbreeding in a dioecious plant on its interaction with pollinating insects and test whether the magnitude of such effects is shaped by plant individual sex and the evolutionary histories of plant populations. We recorded spatial, scent, colour, and rewarding flower traits as well as pollinator visitation rates in experimentally inbred and outbred, male and female Silene latifolia plants from European and North American populations differing in their evolutionary histories. We found that inbreeding specifically impairs spatial flower traits and floral scent. Our results support that sex-specific selection and gene expression may have partially magnified these inbreeding costs for females, and that divergent evolutionary histories altered the genetic architecture underlying inbreeding effects across population origins. Moreover, the results indicate that inbreeding effects on floral scent may have a huge potential to disrupt interactions among plants and nocturnal moth pollinators, which are mediated by elaborate chemical communication.
Keywords: Silene latifolia; dioecy; ecology; evolutionary biology; floral VOC; flower colour; inbreeding depression; sexual dimorphism.
Plain language summary
Destroying habitats can reduce the size of local populations of many plants and animals. For plants, a smaller population means a greater chance of inbreeding, where individual plants that are closely related to each other mate and produce offspring. Inbreeding often results in offspring that are weaker than their parents which can reduce the plant’s chance of survival. Many plants rely on animals to help them to breed. For example, bees carry pollen – containing the male sex cell – to other flowers which then fertilize the plant to produce seeds. Flowers use a wide range of attributes to attract animals such as their colour, scent and providing them with food. However, inbreeding may alter these characteristics which could make it harder for inbred plants to reproduce, meaning that populations would end up shrinking even faster. To test this theory, Schrieber et al. studied flowers from white campions which use moths to breed. Inbred plants had smaller and fewer flowers, and had a different smell. In particular, they produced less of a chemical scent that is known to attract moths at night. Schrieber et al. then tracked moths visiting a mixed population of inbred and control plants. Fewer moths visited the inbred flowers, particularly the ones that were female. This shows that inbreeding may accelerate population loss and extinction by making flowers less attractive to animals. This work highlights the impact habitat destruction has on plants and shows how species can decline rapidly as populations shrink. This could help to support conservation efforts and inform ecology models to better understand our effect on the environment.
© 2021, Schrieber et al.
Conflict of interest statement
KS, LH, AS, RD, JM, CM, AE No competing interests declared
Figures












References
-
- Adams RP. Identification of Essential Oil Components by Gas Chromatography-Mass Spectrometry. 4th Ed. Illinois, United States: Allured Publishing Corporation; 2007.
-
- Angeloni F, Ouborg NJ, Leimu R. Meta-analysis on the association of population size and life history with inbreeding depression in plants. Biological Conservation. 2011;144:35–43. doi: 10.1016/j.biocon.2010.08.016. - DOI
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
