

The bone is the major source of high circulating intact fibroblast growth factor-23 in acute murine polymicrobial sepsis induced by cecum ligation puncture
- PMID: 33989306
- PMCID: PMC8121358
- DOI: 10.1371/journal.pone.0251317
The bone is the major source of high circulating intact fibroblast growth factor-23 in acute murine polymicrobial sepsis induced by cecum ligation puncture
Abstract
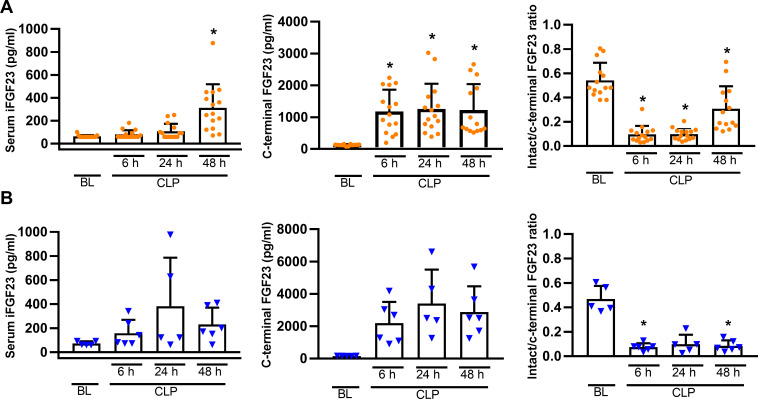
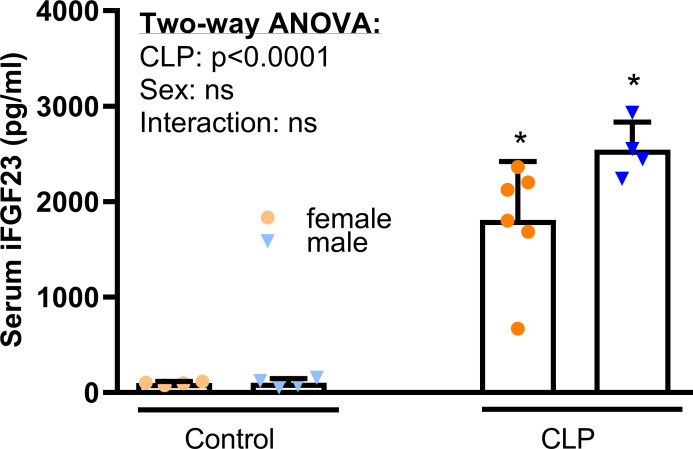
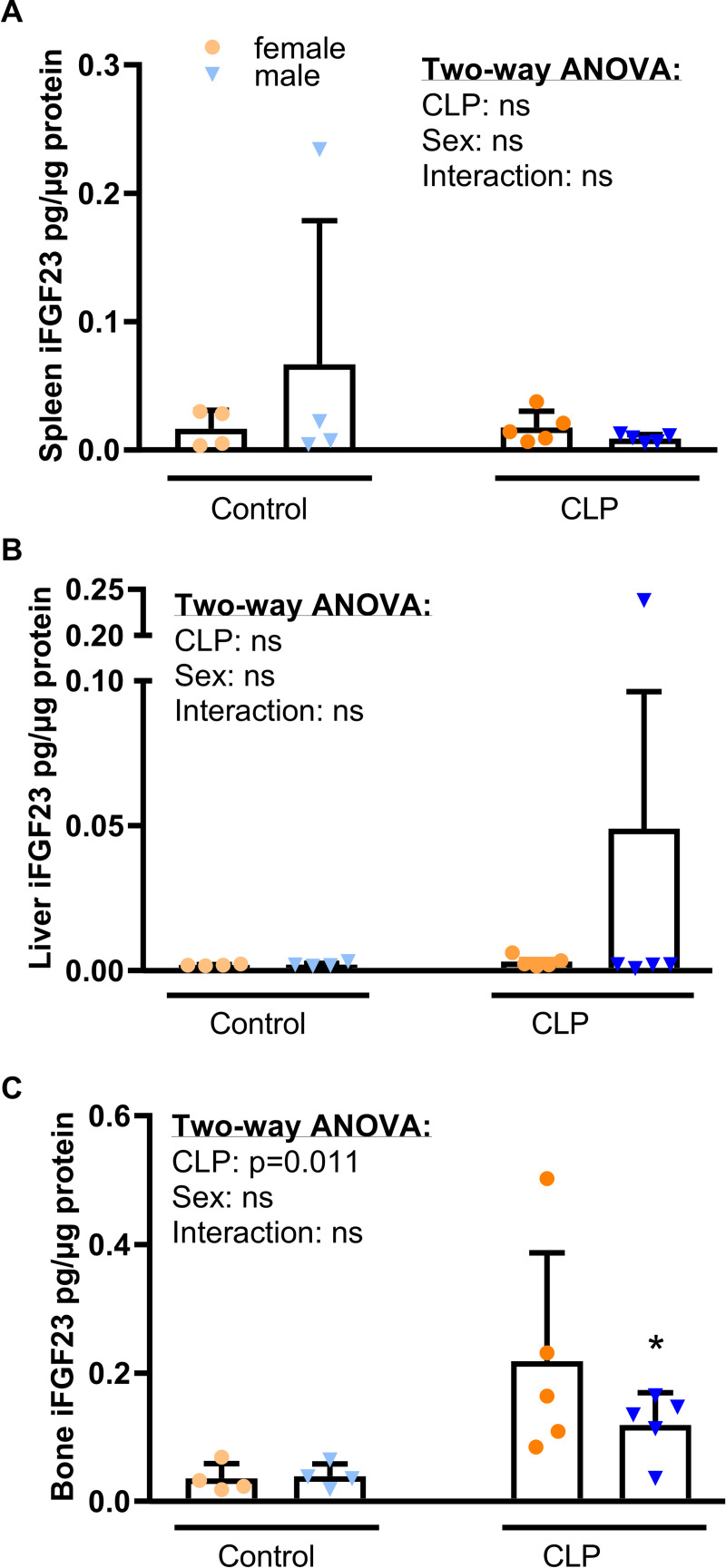
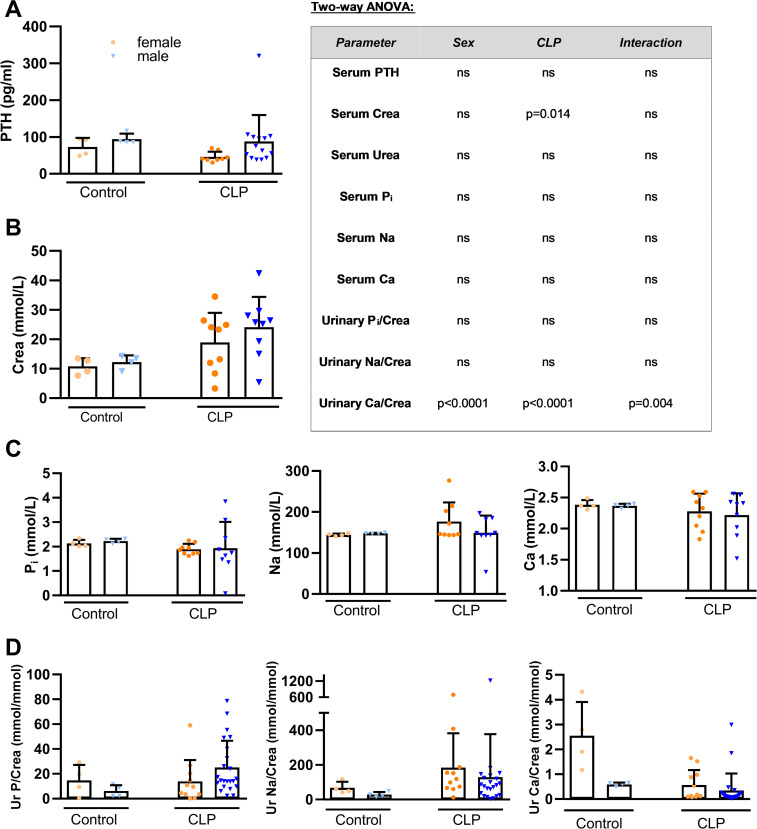
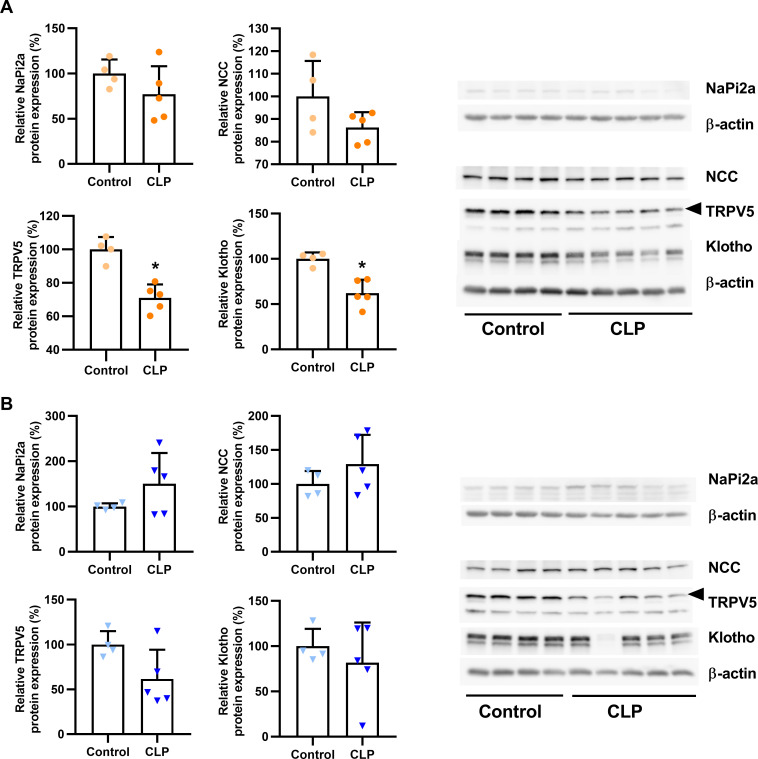
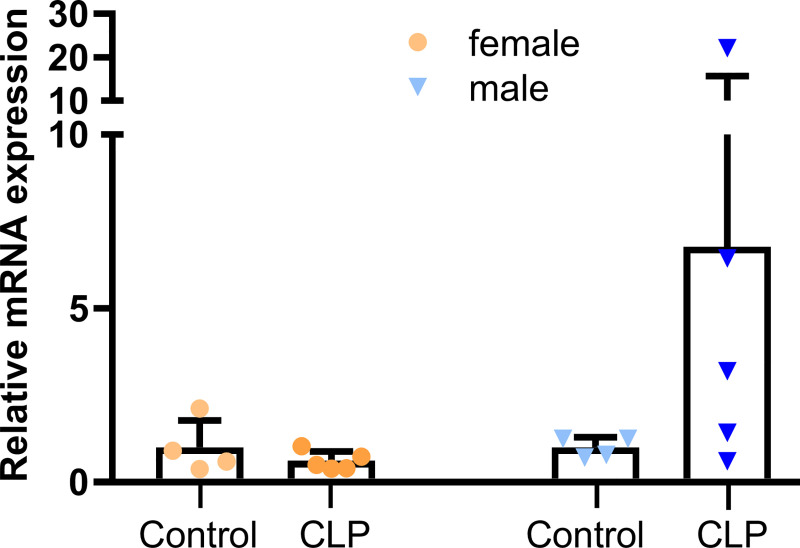
Fibroblast growth factor-23 (FGF23), a bone-produced hormone, plays a critical role in mineral homeostasis. Human diseases associated with excessive intact circulating FGF23 (iFGF23) result in hypophosphatemia and low vitamin D hormone in patients with normal kidney function. In addition, there is accumulating evidence linking FGF23 with inflammation. Based on these studies and the frequent observation of hypophosphatemia among septic patients, we sought to elucidate further the relationship between FGF23 and mineral homeostasis in a clinically relevant murine polymicrobial sepsis model. Medium-severity sepsis was induced by cecum ligation puncture (CLP) in adult CD-1 mice of both sexes. Healthy CD-1 mice (without CLP) were used as controls. Forty-eight hours post-CLP, spontaneous urine was collected, and serum, organs and bones were sampled at necropsy. Serum iFGF23 increased ~20-fold in CLP compared to control mice. FGF23 protein concentration was increased in the bones, but not in spleen or liver of CLP mice. Despite the ~20-fold iFGF23 increase, we did not observe any significant changes in mineral homeostasis or parathyroid hormone levels in the blood of CLP animals. Urinary excretion of phosphate, calcium, and sodium remained unchanged in male CLP mice, whereas female CLP mice exhibited lower urinary calcium excretion, relative to healthy controls. In line with renal FGF23 resistance, expression of phosphate-, calcium- and sodium-transporting proteins did not show consistent changes in the kidneys of male and female CLP mice. Renal expression of the co-receptor αKlotho was downregulated in female, but not in male CLP mice. In conclusion, our data demonstrate that the dramatic, sex-independent rise in serum iFGF23 post-CLP was mainly caused by an upregulation of FGF23 secretion in the bone. Surprisingly, the upsurge in circulating iFGF23 did not alter humoral mineral homeostasis in the acutely septic mice. Hence, the biological function of elevated FGF23 in sepsis remains unclear and warrants further studies.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Effects of iron deficiency anemia and its treatment on fibroblast growth factor 23 and phosphate homeostasis in women.J Bone Miner Res. 2013 Aug;28(8):1793-803. doi: 10.1002/jbmr.1923. J Bone Miner Res. 2013. PMID: 23505057 Clinical Trial.
-
Klotho Lacks an FGF23-Independent Role in Mineral Homeostasis.J Bone Miner Res. 2017 Oct;32(10):2049-2061. doi: 10.1002/jbmr.3195. Epub 2017 Jul 17. J Bone Miner Res. 2017. PMID: 28600880
-
Parathyroid hormone 1 receptor is essential to induce FGF23 production and maintain systemic mineral ion homeostasis.FASEB J. 2016 Jan;30(1):428-40. doi: 10.1096/fj.15-278184. Epub 2015 Oct 1. FASEB J. 2016. PMID: 26428657 Free PMC article.
-
The role of fibroblast growth factor 23 in regulation of phosphate balance.Pediatr Nephrol. 2024 Dec;39(12):3439-3451. doi: 10.1007/s00467-024-06395-5. Epub 2024 Jun 14. Pediatr Nephrol. 2024. PMID: 38874635 Review.
-
[Fibroblast growth factor 23 mediates the phosphaturic actions of cadmium].Nihon Eiseigaku Zasshi. 2012;67(4):464-71. doi: 10.1265/jjh.67.464. Nihon Eiseigaku Zasshi. 2012. PMID: 23095356 Review. Japanese.
Cited by
-
Bone homeostasis disorders increased the mortality of sepsis patients: A preliminary retrospective cohort study.Front Med (Lausanne). 2022 Dec 1;9:1017411. doi: 10.3389/fmed.2022.1017411. eCollection 2022. Front Med (Lausanne). 2022. PMID: 36530881 Free PMC article.
-
Interleukin-36 is overexpressed in human sepsis and IL-36 receptor deletion aggravates lung injury and mortality through epithelial cells and fibroblasts in experimental murine sepsis.Crit Care. 2023 Dec 13;27(1):490. doi: 10.1186/s13054-023-04777-z. Crit Care. 2023. PMID: 38093296 Free PMC article.
-
Intact FGF23 and Markers of Iron Homeostasis, Inflammation, and Bone Mineral Metabolism in Acute Pediatric Infections.Biology (Basel). 2024 Sep 17;13(9):728. doi: 10.3390/biology13090728. Biology (Basel). 2024. PMID: 39336155 Free PMC article.
-
Network pharmacology-based approach to explore the underlying mechanism of sinomenine on sepsis-induced myocardial injury in rats.Front Pharmacol. 2023 Jun 14;14:1138858. doi: 10.3389/fphar.2023.1138858. eCollection 2023. Front Pharmacol. 2023. PMID: 37388447 Free PMC article.
-
Mouse liver injury induces hepatic macrophage FGF23 production.PLoS One. 2022 Mar 1;17(3):e0264743. doi: 10.1371/journal.pone.0264743. eCollection 2022. PLoS One. 2022. PMID: 35231062 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
